Эндотелиальные прогениторные клетки в развитии и восстановлении церебрального эндотелия (обзор)
Рассмотрены современные представления о роли эндотелиальных клеток в формировании и восстановлении гематоэнцефалического барьера головного мозга. Приведены основные характеристики церебральных эндотелиоцитов, входящих в нейроваскулярную единицу, и эндотелиальных прогениторных клеток, описаны их развитие и миграция в головной мозг, молекулярные механизмы барьерогенеза, в том числе в контексте модуляции межклеточных взаимодействий, а также освещены вопросы применения эндотелиальных прогениторных клеток различного происхождения в моделях гематоэнцефалического барьера in vitro. Обсуждаются HIF-1-зависимые механизмы регуляции функциональной активности эндотелиальных клеток, ангиогенеза, астроцит-эндотелиальных взаимодействий и метаболизма клеток нейроваскулярной единицы. Анализируются возможные механизмы участия этих процессов в формировании эндотелиальной дисфункции как одного из компонентов патогенеза нарушений развития головного мозга, его ишемического повреждения или нейродегенерации, ассоциированной с нарушением ключевых метаболических и транспортных процессов, секреции регуляторных молекул, межклеточного сопряжения в клетках церебрального эндотелия, а также с гибелью эндотелиоцитов и потерей структурно-функциональной целостности гематоэнцефалического барьера. Понимание клеточно-молекулярных механизмов барьерогенеза при развитии головного мозга, восстановлении его после повреждения обеспечит новые возможности фармакотерапии заболеваний центральной нервной системы, обусловленных эндотелиальной дисфункцией.
Гетерогенность популяции клеток эндотелия в организме человека определяет особенности функционирования сосудов в разных тканях и органах. Приобретение эндотелиального фенотипа, специфичного для различных тканей, осуществляется на этапах развития эндотелиальных прогениторных клеток путем сложных процессов, определяющих их дифференцировку и миграцию, причем эти процессы достаточно пластичны и могут изменяться под действием внешних факторов [1]. Эндотелиоциты, которые входят в состав нейроваскулярной единицы (НВЕ) головного мозга, составляющей основу структуры гематоэнцефалического барьера (ГЭБ), в процессе эволюции приобрели свойства, позволяющие эффективно регулировать взаимодействие между кровотоком и центральной нервной системой (ЦНС) [2]. Интересно, что эта барьерная функция не всегда ассоциировалась с клетками эндотелиальной природы: изначально ГЭБ формировался из глиальных клеток [3]. Примечательно, что последовательность событий совершенно иная в онтогенезе — ГЭБ формируется в процессе эмбриогенеза из клеток эндотелия и перицитов, и эти события предшествуют включению астроцитов в состав нейроваскулярной единицы [4].
Свойства, определившие преимущества эндотелиоцитов перед клетками глии в контексте регуляции избирательной проницаемости ГЭБ, связаны с возможностью формирования компактного нефенестрированного монослоя, с наличием так называемых тесных контактов, экспрессией большого числа высокоспециализированных транспортных молекул и белков клеточной адгезии, с низким уровнем трансцитоза [5, 6]. Отсутствие фенестр определяет низкий уровень экспрессии гликопротеина MECA-32 (PVLAP) в эндотелии микрососудов головного мозга. Кроме того, эндотелий в составе нейроваскулярной единицы головного мозга является объектом регуляции со стороны других клеток НВЕ — перицитов, астроцитов и нейронов, а также молекул внеклеточного матрикса [7, 8].
Эндотелиальная дисфункция — один из важных механизмов патогенеза нарушений развития головного мозга, его ишемического повреждения или нейродегенерации, что связано с нарушением ключевых метаболических и транспортных процессов, секреции регуляторных молекул, с нарушением межклеточного сопряжения, структурно-функциональной целостности ГЭБ и гибелью эндотелиоцитов. Известно, что целостность ГЭБ определяется многими механизмами, в числе которых Wnt-сигналинг, действие ретиноевой кислоты, контролируемая экспрессия матриксных металлопротеиназ и аквапоринов [9, 10]. Нарушения структурно-функциональной целостности и патологическая проницаемость ГЭБ развиваются практически при всех видах патологии ЦНС, что определяет не только характер течения, но и эффективность фармакотерапии. Чаще всего механизмы нарушения проницаемости ГЭБ ассоциируют с дисфункцией или повреждением зрелых эндотелиоцитов, однако не менее интересным является вклад эндотелиальных прогениторных клеток в эти события.
Эндотелиальные прогениторные клетки: развитие и миграция в головной мозг
Формирование ГЭБ, барьерогенез, — это процесс, реализуемый в эмбриональном и раннем постнатальном периодах развития, а также после перенесенного повреждения головного мозга [11, 12]. Важным компонентом барьерогенеза является ангиогенез, который во многом определяется активностью, пролиферацией и дифференцировкой эндотелиальных прогениторных клеток, в частности, сосуды микроциркуляции в головном мозге формируются из менингеальных сосудов и далее проникают в паренхиму развивающегося мозга за счет механизма ангиогенеза [13]. Кроме того, признаки активации ангиогенеза в головном мозге регистрируются при развитии феномена нейропластичности и запоминания [14].
У лабораторных животных (мышей, крыс) формирование ГЭБ начинается на 10–17-е сутки эмбрионального развития, причем уже к 12-му дню экспрессируются молекулы транспортеров и тесных контактов (у человека экспрессия маркеров ГЭБ регистрируется с 8-й недели развития, а собственно ангиогенез регистрируется в течение 2–3-й недели постнатального периода) [10]. К 21-му дню пренатального развития грызунов формируется трансэндотелиальное сопротивление, оно прогрессивно увеличивается в течение раннего постнатального периода развития, что соответствует формированию полноценных тесных контактов [12]. Этот факт в сопоставлении с данными о динамике эмбрионального нейрогенеза позволил предположить, что барьерогенез инициируется уже после формирования коммитированных нейрональных клеток-предшественников [15].
Во время внутриутробного развития эндотелиальные клетки формируются из общего предшественника и приобретают фенотипические свойства, характерные для специализированной ткани. В процессе эмбриогенеза эндотелиальные клетки заселяют регионы формирующегося мозга, содержащие нейроэпителиальные клетки, радиальную глию, нейробласты и нейроны [16]. Иммунофенотипирование эндотелиальных прогениторных клеток демонстрирует экспрессию ими некоторых ключевых антигенов (например, CD34, CD31, CD133, CD45, VEGFR2, CD144 и др.) [17], идентификация которых на резидентных клетках сосудистой стенки или прогениторных клетках костно-мозгового происхождения позволяет судить о механизмах развития и направленной миграции клеток эндотелия в ткани.
Эмбриональные стволовые клетки, коммитированные по эндотелиальному пути развития, рано начинают экспрессировать молекулы межклеточных контактов (молекулы клеточной адгезии, белки тесных и щелевых контактов) [18]. Вместе с тем, согласно экспериментальным данным [19], эндотелиальные прогениторные клетки, циркулирующие в периферической крови, способны приобретать фенотип, специфичный для ГЭБ, уже находясь в специализированных условиях культивирования in vitro, что необходимо учитывать при разработке моделей ГЭБ.
Молекулярные механизмы направленной миграции эндотелиальных прогениторных клеток в ткань мозга изучены недостаточно. Вероятнее всего, мобилизация и рекрутинг эндотелиальных прогениторных клеток в развивающемся и зрелом мозге осуществляются благодаря продукции паракринных и эндокринных сигналов, продуцируемых клетками НВЕ в (пато)физиологических условиях.
Клетки радиальной глии и нейроэпителиальные клетки первыми взаимодействуют с эндотелиальными клетками в головном мозге в эмбриогенезе. Клетки радиальной глии экспрессируют глутамат-аспартатный транспортер GLAST, что указывает на их роль в регуляции метаболических событий на ранних этапах развития клеток головного мозга, а также коннексин 43 (Сх43), обеспечивающий эффективную паракринную и аутокринную сигнализацию в участках интенсивной пролиферации клеток [20]. Именно клетки радиальной глии продуцируют ретиноевую кислоту, являющуюся одним из наиболее эффективных регуляторов формирования ГЭБ [9]. Применительно к процессам нейрогенеза полагают, что в зрелом мозге часть клеток, имеющих фенотип радиальной глии, наиболее активной в периоде эмбрионального нейрогенеза, сохраняется в нейрогенных нишах, не превращаясь в астроциты, что обеспечивает репаративный нейрогенез [21]. YKL-иммунопозитивные клетки, относящиеся к клеткам-предшественникам астроглии, обнаруживаются в головном мозге в раннем периоде развития именно в участках интенсивного ангиогенеза и формирующегося ГЭБ [22]. Интересно, что астроциты, экспрессирующие YKL-40, маркируют участки нейровоспаления или нейродегенерации в зрелом мозге [23, 24], что может свидетельствовать о роли астроглии в регуляции репаративного ангиогенеза. В периоде эмбриогенеза и в зрелом мозге эндотелиальные клетки сохраняют за собой важную функцию регуляторов нейрогенеза, что подтверждается их стимулирующим влиянием на поддержание популяции стволовых и прогениторных клеток головного мозга, а также действием факторов роста (например, VEGF, BDNF, LIF, PDGF, IGF) и других регуляторных молекул эндотелиального происхождения на клетки-участники процесса нейрогенеза [25, 26].
В постнатальном периоде неоваскулогенез, например при повреждении головного мозга, обеспечивается эндотелиальными прогениторными клетками костно-мозгового происхождения [27], и, вероятнее всего, именно этот механизм актуален для репарации ГЭБ, что объясняет зависимость клинического эффекта после повреждения головного мозга от уровня мобилизации эндотелиальных прогениторных клеток в организме [28]. Пролиферация эндотелиальных клеток может индуцироваться, например, гипоксией, после чего активируются астроциты, изменяя экспрессию молекул клеточной адгезии в местах контакта с клетками эндотелия [29].
Нарушение различных этапов барьерогенеза и функционирования ГЭБ индуцируется многими факторами: 1) подавлением во время внутриутробного периода васкулогенеза и нейрогенеза; 2) влиянием патологии беременности и перинатального стресса на регуляцию развития и функционирования ГЭБ и на транспортные функции клеток эндотелия гуморальными факторами (гормонами стресса, нейропептидами, интерлейкинами); 3) повреждением нейронов, активацией астроцитов и микроглии, развитием нейровоспаления при нейроинфекциях и ишемии, что вызывает повышение проницаемости ГЭБ, особенно очевидное у недоношенных детей; 4) системным воспалением и гиперпродукцией провоспалительных цитокинов в пренатальном и раннем постнатальном периодах, которые вызывают раннее повреждение ГЭБ и нарушение васкулогенеза с последующим формированием патологической проницаемости ГЭБ во взрослом периоде развития организма [30, 31].
Молекулярные механизмы барьерогенеза: роль межклеточных взаимодействий
Механизмам барьерогенеза посвящено много интересных обзоров литературы [10, 12, 32 и др.]. Согласно современным представлениям, сложные процессы барьерогенеза регулируются большим количеством факторов, в числе которых ведущая роль отводится Wnt/catenin, сиртуинам, Notch, HIF-1, NF-kB, FOX, RhoАТФ-азе, GSK-3, а также гуморальным регуляторам (сосудисто-эндотелиальный фактор роста (VEGF), матриксные металлопротеиназы, нейротрансмиттеры, нейропептиды, нейростероиды, ФНО-α, ИЛ-1) [12, 32, 33]. На всех этапах барьерогенеза важным является формирование локального микроокружения (как правило, поддерживаемого гуморальными регуляторами), обеспечивающего разнообразные межклеточные взаимодействия, которые контролируют пролиферацию и дифференцировку клеток [34]. Существенную роль в формировании соответствующего локального микроокружения играют взаимодействия соседствующих (в пределах нейроваскулярной единицы) клеток — эндотелиоцитов, астроцитов, перицитов, нейронов. Интересно, что такие взаимодействия актуальны для всех этапов онтогенеза, и совершенно очевидно, что их характер меняется в процессе развития ГЭБ, на фоне его повреждения или репарации. Так, показано, что совместное культивирование in vitro эмбриональных нейрональных клеток-предшественников с эндотелиальными клетками церебральных капилляров индуцирует в последних барьерные свойства, характерные для эндотелия ГЭБ [35]. Позднее возможность воспроизведения сочетанных механизмов нейрогенеза и ангиогенеза из стволовых клеток in vivo была продемонстрирована M. Ii с соавт. [36].
Индукция фенотипа, характерного для клеток ГЭБ, в эндотелиоцитах в период пренатального развития головного мозга обеспечивается влиянием на эндотелий стволовых нейрональных клеток [4].
Важную роль в ангиогенезе в развивающемся и зрелом мозге играют перициты. Эти клетки, участвующие в формировании ГЭБ, являются объектом регуляторного влияния большого числа гуморальных факторов эндотелиального происхождения, в частности VEGF и ангиопоэтинов [37–39]. Интересно, что разные сигнальные пути вовлечены в механизмы участия перицитов в формировании сосудов и барьерогенезе [40].
В целом в процессе барьерогенеза фактически можно наблюдать три основных направления специализации клеток-предшественников: 1) нейронализация (Wnt/Notch-регулируемые и другие ассоциированные механизмы); 2) астроглиализация (STAT3/RAR/Zac1-регулируемые и другие ассоциированные механизмы); 3) эндотелиолизация (HIF-1/SIRT1-регулируемые и другие ассоциированные механизмы).
Несмотря на то, что зрелые астроциты не принимают участия в ранних этапах формирования ГЭБ (в окружении мигрирующих в мозг клеток эндотелия преимущественно обнаруживаются клетки радиальной глии), существует точка зрения, что астроциты обеспечивают формирование «матрицы» для ангиогенеза в процессе развития головного мозга за счет продукции VEGF, находясь в тесном взаимодействии с прогениторными клетками, формирующими сосуды [41]. Кроме того, есть данные о том, что метаболическая дисфункция астроцитов определяет характер повреждения ГЭБ [42]. Ретиноевая кислота — важный регулятор барьерогенеза [9] — является продуктом активности астроцитов [43]. Реактивные астроциты регулируют взаимодействие клеток церебрального эндотелия и эндотелиальных прогениторных клеток, высвобождая во внеклеточную среду белки группы HMGB1 (например, как результат повреждения клетки и активации инфламмасом), далее взаимодействующие с RAGE-рецепторами на клетках эндотелиальной природы [44].
HIF-1-сопряженные механизмы в астроцитах и эндотелиоцитах контролируют процессы ангиогенеза и барьерогенеза, проницаемости ГЭБ [45]; действие нейротоксических факторов, в частности амилоида, ассоциировано с повреждением астроцитов и вызывает интенсивный ангиогенез, нарушает проницаемость ГЭБ [46]. Такие пептиды астроглиального происхождения, как VEGF, стимулируют ангиогенез, тогда как тромбоспондин обладает ангиостатическим эффектом, но они оба важны для репаративного барьерогенеза и нейрогенеза [47, 48]. Интересно, что гиппокампальные механизмы пластичности определяются особенностями продукции тромбоспондина астроцитами [49], а ангиогенный потенциал эндотелиальных прогениторных клеток подавляется тромбоспондином-1 [50]. Факторы, оказывающие влияние на биологические эффекты этих молекул (матриксные металлопротеиназы MMPи их ингибиторы TIMP, рецепторы тромбоспондинов CD47 и CD36), могут рассматриваться в качестве молекул-участников патогенеза большого спектра заболеваний [51].
Одним из самых мощных факторов стимуляции ангиогенеза, в частности при патологических состояниях, ассоциированных с необходимостью дополнительной васкуляризации ткани или восстановления поврежденных сосудов, является гипоксия. Интересно, что гипоксия повышает экспрессию PVLAP на клетках эндотелия ГЭБ, что может свидетельствовать об увеличении степени фенестрации и уровня трансцитоза [4]. При гипоксии интересной является пока еще мало изученная функция представителей семейства Rho ГТФ-азы, например Rac1, под контролем которого находятся следующие события в клетках различной природы: 1) экспрессия HIF-1, определяющая характер ответа клеток на гипоксию, эффекты инсулина и метаболизм глюкозы [52]; 2) изменения белков цитоскелета, в частности необходимые для так называемой стеллации астроцитов [53]; 3) продукция свободных радикалов, секреция матриксных протеиназ (MMP-2, MMP-9), что является важным для ремоделирования внеклеточного матрикса [54]. Очевидно, что все перечисленные механизмы актуальны как для функционирования ГЭБ, так и для процессов барьерогенеза. Действительно, поддержание структурно-функциональной целостности эндотелиального барьера нуждается в активности Rac1 в различных органах и тканях [55–57]. Миграция эндотелиальных клеток в специализированные ткани требует участия Rac1 в реализации эффектов SDF-1 (stromalcell-derivedfactor-1), что актуально для ангиогенеза [58], например при повреждении ткани [59].
Многие события при межклеточных взаимодействиях определяются активностью транскрипционного фактора HIF-1, опосредующего ответ клеток на гипоксию. Известно, что HIF-1-контролируемые реакции энергетического обмена находят свое отражение в изменении процессов гликолиза, аккумуляции лактата и изменении характера нейрон-астроглиального метаболического сопряжения. Активность HIF-1 регулирует экспрессию VEGF [60] и церебральный ангиогенез. Кроме того, в числе HIF-1-контролируемых генов — гены, кодирующие SDF-1, транспортеры глюкозы и лактата, ферменты гликолиза, что необходимо для обеспечения функционирования клеток в условиях острой или хронической гипоксии. Интересно, что продукт анаэробного гликолиза — лактат (ключевой регулятор глиоваскулярного контроля, обеспечивающего увеличение локального кровотока в функционально активных регионах мозга) — способствует реализации программы ангиогенеза, действуя на клетки эндотелия [61], стимулирует циркулирующие стволовые васкулогенные клетки и увеличение экспрессии в них HIF-1-контролируемых ангиогенных факторов роста [62]. Проангиогенная активность лактата очевидна в условиях, при которых добавление лактата в матрицу для развития клеток сосудов существенно улучшает параметры ангиогенеза в зоне размещения имплантата [63].
Активация HIF-1 в клетках может быть индуцирована как гипоксией, так и негипоксическими стимулами, например продукцией активных форм кислорода митохондриями [64]. Важно отметить, что продукция лактата тесно связана с редокс-состоянием клетки [65], в частности соотношением НАД/НАДН в митохондриях, поэтому негипоксические механизмы активации HIF-1 могут быть особенно актуальными в физиологических условиях функционирования ГЭБ. Насколько важны в этом контексте митохондриальные события в клетках эндотелия, сказать трудно, особенно с учетом особенностей функционирования митохондрий в клетках эндотелиальной природы по сравнению с другими клетками [66].
В пределах гематоэнцефалического барьера основными продуцентами лактата являются астроциты и клетки эндотелия. Так, показано, что продукция NO эндотелиоцитами стимулирует HIF-1α-зависимые эффекты (в частности, активацию анаэробного гликолиза) в контактирующих астроцитах [67]. С другой стороны, пролиферация клеток эндотелия сама по себе контролируется процессами метаболизма глюкозы, а продуцируемый в ткани лактат транспортируется в клетки эндотелия, вызывая в них HIF-1α-контролируемые события, приводящие к стимуляции ангиогенеза [68], что было продемонстрировано в опухолевой ткани. Насколько аналогичный механизм справедлив для астроцит-эндотелиальных взаимодействий в церебральных сосудах, нуждается в экспериментальной оценке, однако с учетом высокой гликолитической активности астроцитов по сравнению с другими клетками нейроваскулярной единицы и их контролирующего влияния на процессы метаболического сопряжения клеток этот механизм представляется весьма вероятным.
Интересно, что ответ астроцитов на гипоксию, связанный с активацией HIF-1, имеет значение для патологического ангиогенеза, в свою очередь связанного с повреждением клеток центральной нервной системы, но не оказывает существенного влияния на ангиогенез в процессе развития мозга [69]. Согласно данным другой исследовательской группы, именно астроцитам принадлежит ключевая роль в формировании ангиогенного микроокружения при ангиогенезе, но HIF-1-α-опосредованные механизмы при этом локализованы преимущественно в нейронах, контактирующих с астроцитами [41].
В центральной нервной системе HIF-1 влияет на процессы транспорта и захвата глюкозы клетками, запоминания [70], а лактат выступает в качестве глиотрансмиттера, контролируя возбудимость нейронов [71], и в качестве ключевого регулятора локального кровотока в активных зонах мозга. Одновременно HIF-1 является объектом эпигенетической регуляции в клетках нейроваскулярной единицы, будучи деацетилированным сиртуином 1 [72], что может иметь отношение как к эпигенетической регуляции когнитивных функций в развивающемся мозге [73], так и к гомеостазу НАД+, определяющему активность НАД+-зависимых ферментов (АДФ-рибозилполимераза, АДФ-рибозилциклаза/CD38/CD157, сиртуины) [74]. Интересно, что лактат как продукт гликолитической активности подавляет активность деацетилаз (сиртуинов), вызывая долгосрочные изменения экспрессии генов [75]. Существует взаимосвязь между интенсивностью гликолиза и синаптогенеза, а также экспрессией генов, характерных для ранних этапов развития мозга [76]; между экспрессией генов, кодирующих белки, вовлеченные в нейрон-астроглиальное метаболическое сопряжение, и состояниями, связанными с нарушением клиренса метаболитов через гематоэнцефалический барьер [77].
Разработка современных фармакотерапевтических стратегий, направленных на регуляцию HIF-1-регулируемых механизмов ангиогенеза, является перспективным направлением в повышении эффективности лечения ишемических повреждений и дегенерации тканей, контроля опухолевой прогрессии [78], однако к настоящему времени есть лишь единичные доказательства того, что такие стратегии оправдают себя при коррекции нарушений барьерогенеза или повреждения ГЭБ [79–81].
Применение эндотелиальных прогениторных клеток в моделях гематоэнцефалического барьера in vitro
Одна из ключевых задач при моделировании ГЭБ in vitro — получение клеток-компонентов системы, наиболее близких по фенотипическим свойствам к клеткам ГЭБ in vivo (межклеточные контакты, метаболизм, активность белков-транспортеров, процессы эндо- и экзоцитоза), а также воспроизведение с максимальной точностью естественных межклеточных взаимодействий, составляющих основу барьера [82]. Наиболее критическим моментом является использование максимально релевантных по отношению к ситуации in vivo (в контексте реализации собственно барьерной функции) клеток эндотелия. Другой критический момент — воспроизведение адекватной секреторной активности клеток-компонентов ГЭБ, обеспечивающих межклеточные взаимодействия и поддержание целостности барьера. Так, значительное число гуморальных факторов эндотелиальной природы и белков базальной мембраны ГЭБ влияет на состояние астроцитов, входящих в состав НВЕ [83].
Для решения этих задач предложены несколько подходов (см. таблицу). Наиболее часто используют метод, базирующийся на получении зрелых нейронов, астроцитов, эндотелиоцитов, выделенных из капилляров мозга, перицитов. Вместе с тем в течение последних трех лет сделаны попытки получения клеток-компонентов ГЭБ и их полной характеристики (в том числе с применением методов метаболомики, протеомики) из клеток-предшественников (прогениторные клетки головного мозга) [84]. Такой подход интересен тем, что возможны и воспроизведение естественных механизмов так называемого барьерогенеза in vitro, представляющего по своей сути сочетание нейрогенеза и ангиогенеза, и идентификация ключевых клеточных событий, определяющих функциональную компетентность ГЭБ. В частности, анализ секретома эндотелиальных прогениторных клеток позволил идентифицировать молекулярные механизмы их влияния на ангиогенную активность церебрального эндотелия [27].
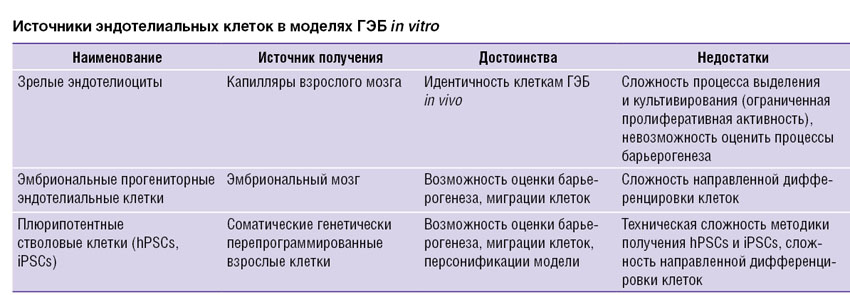 Источники эндотелиальных клеток в моделях ГЭБ in vitro Источники эндотелиальных клеток в моделях ГЭБ in vitro
|
В настоящее время в качестве источника получения нейронов и астроцитов в таких протоколах применяются попытки использования нейрональных прогениторных клеток (NPCs) [85], а для получения клеток эндотелия — дифференцирующиеся человеческие плюрипотентные стволовые клетки (hPSCs). Интересно, что около 6% нейрональных стволовых клеток, культивируемых в присутствии эндотелия, могут конвертироваться в клетки эндотелиальной природы [86], что открывает возможности направленной дифференцировки нейрональных стволовых клеток in vitro во все основные клеточные компоненты ГЭБ (нейроны, астроциты, эндотелиоциты). Вместе с тем с учетом выраженной гетерогенности популяции нейрональных стволовых клеток подбор адекватных условий культивирования для достижения указанного результата представляет собой серьезную методологическую проблему.
Получение клеток эндотелия, имеющего характеристики церебрального эндотелия, из hPSCs поможет осуществить настоящий прорыв в моделировании ГЭБ для решения задач персонифицированного скрининга нейротропных лекарственных препаратов, коль скоро все существующие на сегодняшний день модели ГЭБ утилизируют либо эндотелиоциты, выделенные из капилляров мозга (и это делает практически невозможным создание абсолютно адекватной модели ГЭБ, характерной для конкретного пациента), либо эндотелиоциты клеточных линий немозгового происхождения, например HUVEC (что ставит под сомнение возможность полной экстраполяции полученных данных на барьерные структуры центральной нервной системы).
Применение в среде культивирования гуморальных факторов, актуальных для барьерогенеза in vivo, например ретиноевой кислоты [87], позволяет существенно улучшить результаты получения клеток эндотелия, максимально приближенных по своим свойствам к церебральному фенотипу.
Еще один методический подход заключается в получении эндотелиальных клеток-компонентов ГЭБ из гемопоэтических стволовых клеток пуповинной крови, которые в соответствующих условиях начинают экспрессировать белки тесных контактов и транспортеры, характерные для клеток церебрального эндотелия в составе ГЭБ [88].
Использование hPSCs в качестве источника получения так называемого примитивного эндотелия (эндотелия, характерного для ранних этапов онтогенеза) ставит дополнительную задачу по поиску оптимальных условий сокультивирования нейронов, астроцитов (полученных из плюрипотентных или прогениторных клеток) и эндотелиальных клеток для достижения последними максимально зрелого фенотипа (характерного для зрелого мозга) [16].
Разработка протоколов получения индуцированных человеческих плюрипотентных клеток (iPSCs) открыла новую эру в экспериментальной неврологии и нейрофармакологии [89–91]. Применение iPSCs для моделирования in vitro структур мозга, в том числе ГЭБ, открывает новые возможности в разработке технологий персонифицированного скрининга лекарственных веществ, а также в изучении индивидуальных особенностей патогенеза заболеваний центральной нервной системы.
Клеточно-молекулярные механизмы барьерогенеза и поддержания целостности ГЭБ в пре- и постнатальном периодах развития головного мозга отличаются друг от друга, что определяется не только особенностями функционирования эндотелиальных прогениторных клеток, но и их взаимодействиями с другими клетками НВЕ. Прогресс в изучении этих механизмов позволит разработать новые фармакотерапевтические подходы к направленной регуляции проницаемости ГЭБ.
Финансирование исследования. Исследование выполнено за счет гранта Российского научного фонда (проект №14-25-00054).
Конфликт интересов. У авторов нет конфликта интересов.








