Роль генетических факторов человека в естественной селекции доминирующего генотипа вируса гепатита C в этнически близких популяциях бурят и халха-монголов
Цель исследования — определить роль полиморфизма генов врожденного иммунитета в популяционной селекции генотипов вируса гепатита С (HCV), циркулирующих в этнически близких группах монголоидов: бурят и халха-монголов.
Материалы и методы. Генетическое определение нуклеотидных полиморфизмов генов врожденного иммунитета выполнено у 400 больных хроническим гепатитом С, из них 200 человек принадлежат к этнической группе монголов, проживающих в Улан-Баторе (Монголия), и 200 человек — к этнической группе бурят, проживающих в Улан-Удэ (Республика Бурятия). Контрольную группу (n=531) составили практически здоровые лица, из которых 220 были бурятами, а 311 — халха-монголами. В указанных выборках больных и здоровых лиц выполнены генетические исследования двенадцати однонуклеотидных полиморфизмов девяти генов: IFNL1 (rs30461); IFNL3 (rs12979860 и rs8099917); IFNL4 (rs368234815); CD209 (rs4804803); TLR3 (rs3775291 и rs13126816); TLR7 (rs179008 и rs179009); IFITM (rs12252); MyD88 (rs6853); IFIH1 (rs1990760). При анализе результатов генетических исследований проводили сравнение показателей встречаемости аллелей генов и их комбинаций в виде генотипов.
Результаты. На территории Монголии обнаружено доминирующее преобладание 1-го генотипа HCV (98,0%), что значимо выше (р<0,001) его распространенности на территории Бурятии (66,0%). Среди генетических факторов, которые могут оказывать влияние на формирование структуры циркулирующих генотипов в популяции бурят и монголов, выявлены однонуклеотидные полиморфизмы трех генов (IFNL3, TLR3 и TLR7), частота встречаемости которых в изучаемых когортах существенно отличалась. В этнической группе бурят поиск кандидатных генов у больных хроническим гепатитом C при 1-м и не 1-м (2-й или 3-й, 2/3) генотипах вируса установил, что Т-аллель rs179008 гена TLR7 более чем в 2 раза чаще встречается у женщин, больных хроническим гепатитом C, вызванным 2/3-м генотипом, чем 1-м (р=0,04).
Заключение. Низкая распространенность 2-го и 3-го генотипов HCV на территории Монголии может быть обусловлена редкой встречаемостью среди населения мутантного Т-аллеля гена TLR7 (rs179008), ассоциированного с предрасположенностью к инфицированию HCV-2/3, что в настоящей работе продемонстрировано на примере этнической группы бурят.
Введение
Разработка комплекса мер профилактики и внедрение в клиническую практику препаратов прямого противовирусного действия привели к стабилизации уровня заболеваемости хроническим вирусным гепатитом С (ХГС). В настоящее время, по данным ВОЗ, количество больных ХГС существенно сократилось — со 160 до 71 млн. человек [1, 2]. В 2016 г. на 69-й Всемирной ассамблее здравоохранения была принята резолюция, в которой продекларировано существенное снижение заболеваемости вирусным гепатитом в мире к 2030 г. вплоть до его полной элиминации. Необходимо учитывать, что в странах Азиатско-Тихоокеанского региона сосредоточено более 50% всех инфицированных на планете [1]. В Азии наблюдаются самые высокие показатели заболеваемости гепатоцеллюлярной карциномой и частоты формирования цирроза печени в исходе вирусных гепатитов [1].
Вирус гепатита С (HCV) отличается высокой изменчивостью и широким генотипическим разнообразием. Определенные генотипы HCV превалируют в отдельных географических регионах. Например, генотип 1 (HCV-1) доминирует в странах Южной и Северной Америк, Австралии и Европе (53–71%), в то время как генотип 3 (HCV-3) наиболее распространен в Азии (40%), а генотип 4 (HCV-4) — в Северной Африке и на Ближнем Востоке (около 71%) [3]. Несмотря на большой объем информации, связанной с распространением генотипов HCV в конкретных странах, до конца неясен механизм формирования стабильных эпидемических генотипов в ограниченных географических ареалах.
В Монголии отмечается один из наиболее высоких показателей заболеваемости вирусным гепатитом C в Азии, при этом превалирующее распространение среди всех генотипов имеет HCV-1b (98,8%) [4, 5]. Феномен абсолютного доминирования HCV-1 не наблюдается ни в соседней Бурятии, ни в Китае, ни в других азиатских странах [3]. Такую высокую концентрацию одного генотипа в популяции жителей Монголии достаточно сложно объяснить особенностями эпидемиологии вирусного гепатита C в этой стране, или «эффектом основателя». При этом в России, в том числе в этнически близкой Бурятии, доля HCV-1b не превышает 57,1% [6], а в Китае — 56,8% [3].
Одним из вероятных объяснений имеющегося феномена может быть наличие определенной толерантности населения к заражению HCV-2/3, при котором HCV-1 получает селективное преимущество. Можно предположить, что вышеуказанная предрасположенность имеет генетическую природу и может быть обнаружена при сравнительном исследовании генетического полиморфизма этнически близких популяций монголов и бурят. Важным для объяснения причин естественной селекции доминирующего генотипа HCV на территории Монголии может стать исследование полиморфизма генов врожденного иммунитета, кодирующих синтез мембранных и внутриклеточных адаптерных молекул — эффекторов иммунного ответа на HCV [7–9].
Цель исследования — определить роль полиморфизма генов врожденного иммунитета в популяционной селекции генотипов вируса гепатита С, циркулирующих в этнически близких группах монголоидов: бурят и халха-монголов.
Материалы и методы
Представленные исследования проводили с 2013 по 2017 г. на территории России (Республика Бурятия) и Монголии. Сбор материала осуществляли в лечебно-профилактических учреждениях Бурятии и Монголии: Республиканской клинической инфекционной больнице (Улан-Удэ, Республика Бурятия) и Национальном центре по изучению инфекционных болезней (Улан-Батор, Монголия).
Генетическое определение однонуклеотидных полиморфизмов (ОНП) генов врожденного иммунитета выполнено у 400 больных ХГС, среди которых 200 человек принадлежат к этнической группе халха-монголов, проживающих в Улан-Баторе, и 200 человек — к этнической группе бурят, проживающих в Улан-Удэ. Среди представителей монголоидной расы в Монголии наиболее многочисленной является этническая группа халха-монголов, которая составляет более 85% от общей численности населения [10].
На обеих территориях больных отбирали сплошным случайным методом из всех обратившихся в Гепатологический центр инфекционной больницы Улан-Удэ (Республика Бурятия) и Центр по изучению инфекционных заболеваний Улан-Батора (Монголия). Диагноз ХГС устанавливали на основании данных анамнеза, клинического обследования, определения активности печеночных трансаминаз, выявления анти-HCV IgG и РНК HCV. Контрольную группу (n=531) составили практически здоровые лица, из которых 220 были бурятами, а 311 — халха-монголами.
Генетическое исследование проведено в соответствии с Хельсинкской декларацией (2013) и одобрено Этическим комитетом Иркутского государственного медицинского университета. Все обследуемые лица дали информированное согласие на его проведение.
Генотипирование вируса методом ПЦР осуществляли с использованием набора «АмплиСенс-НСV-генотип» (Центральный НИИ эпидемиологии Роспотребнадзора РФ).
С целью оценки возможного влияния генетических маркеров на селекцию определенного генотипа вируса, доминирующего в популяции, в указанных выборках больных и здоровых лиц выполнены генетические исследования двенадцати ОНП девяти генов: IFNL1 (rs30461); IFNL3 (rs12979860 и rs8099917); IFNL4 (rs368234815); CD209 (rs4804803); TLR3 (rs3775291 и rs13126816); TLR7 (rs179008 и rs179009); IFITM (rs12252); MyD88 (rs6853); IFIH1 (rs1990760). При выборе ОНП авторы рассматривали полиморфизмы в генах, которые кодируют синтез рецепторов или адаптерных молекул, имеющих большое значение в качестве противовирусных факторов врожденного иммунитета [11]. Предварительный анализ источников литературы [7–9, 11, 12] показал наличие сигнальных белков, чьи гены практически не изучались при вирусном гепатите С, но которые играют большую роль в обеспечении врожденной устойчивости при других вирусных инфекциях.
Молекулярно-генетические исследования осуществляли с использованием амплификатора для проведения ПЦР в режиме реального времени Rotor-Gene Q (Qiagen Hilden, Германия). Амплификацию определенных участков генов выполняли с помощью соответствующих праймеров. Анализ генотипов гена IFNL3 проводили с помощью набора реагентов для определения ОНП rs8099917 и rs12979860 «АмплиСенс» (Центральный НИИ эпидемиологии Роспотребнадзора РФ) методом ПЦР-РВ. Для определения остальных генотипов использовали праймеры и зонды собственного дизайна (табл. 1), синтезированные в НПФ «Синтол» и ЗАО «Евроген» (Россия).
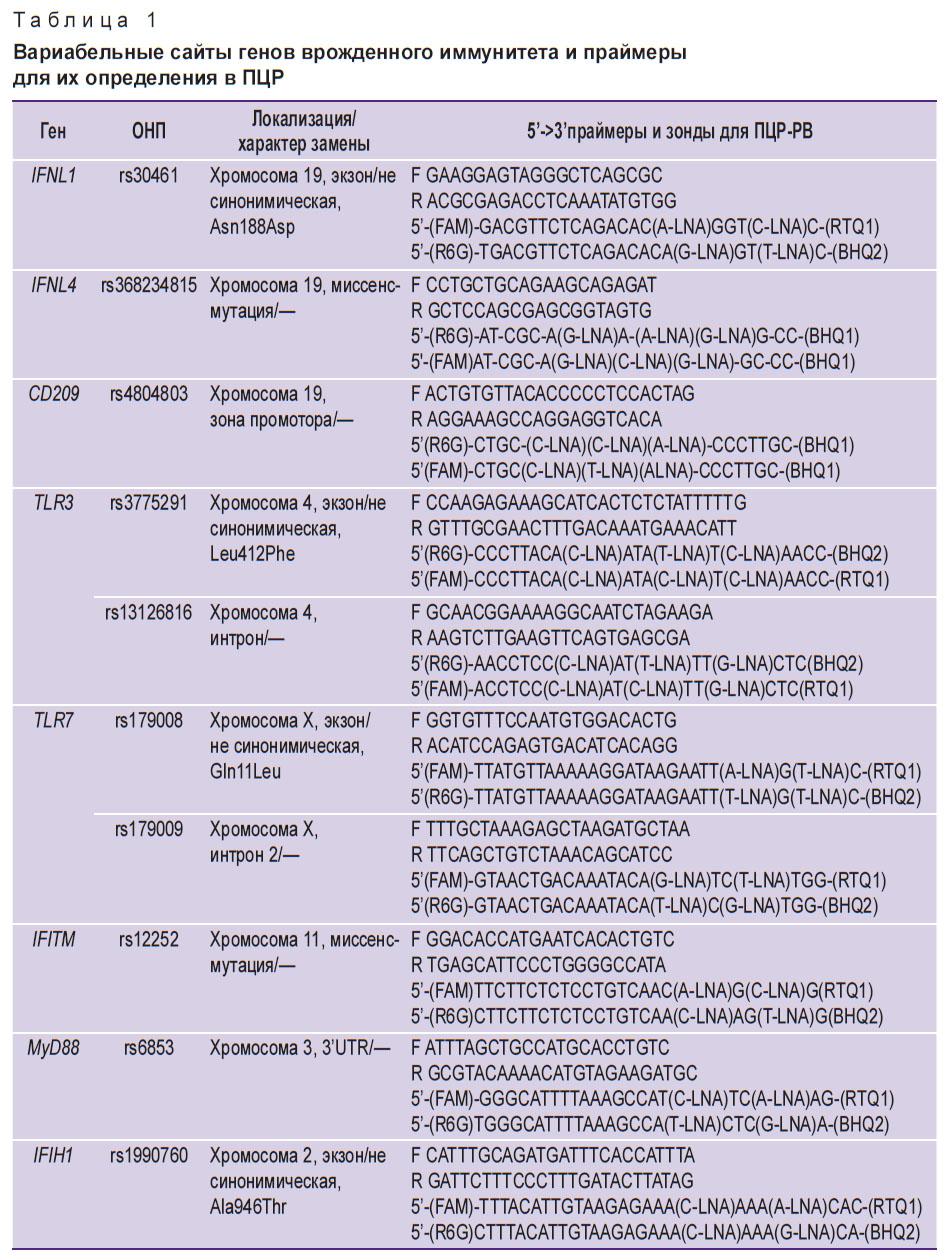
|
Таблица 1. Вариабельные сайты генов врожденного иммунитета и праймеры для их определения в ПЦР |
Для повышения эффективности амплификации использовали технику «горячего старта» с полимеразой TaqF («Интерлабсервис», Россия). Режим ПЦР: денатурация пробы при температуре 95°С в течение 15 мин, а затем 45 циклов: 95°С — 15 с, 60°С — 30 с.
Статистическую обработку первичных материалов проводили с использованием общепринятых параметрических и непараметрических критериев статистики в программе Statistica 6.1 (StatSoft Inc., США). Для статистического анализа кандидатных генетических исследований использовали программу SNPStats (http://bioinfo.iconcologia.net/SNPStats). Для каждого ОНП статистический анализ был выполнен по различным моделям наследования: кодоминантной, доминантной, рецессивной, сверхдоминантной и лог-аддитивной. При этом модель с наименьшим значением информационного критерия Акаике (Akaike information criterion) для каждого генного полиморфизма принималась в качестве наиболее вероятной [13]. При анализе результатов генетических исследований проводили сравнение показателей встречаемости аллелей генов и их комбинаций в виде генотипов. Распределение генотипов по каждому локусу проверяли на соответствие закону Харди–Вайнберга. Расчет отношения шансов (OR — оdds ratio) проводили при уровне доверительного интервала 95% CI (95% confidence interval). Расчет величины OR при нулевом значении показателя осуществляли по методу Вульфа–Холдейна [14]. Уровень статистической значимости принят при р≤0,05.
Результаты
Сравнительный анализ клинико-лабораторных показателей у больных ХГС монголов и бурят приведен в табл. 2.

|
Таблица 2. Клинико-лабораторная характеристика больных хроническим гепатитом С в этнических группах бурят и монголов (M±m) |
Клиническая симптоматика не показала каких-либо особенностей течения ХГС. Единственное статистически значимое отличие такого показателя, как уровень гемоглобина в крови, отражает физиологическую реакцию организма на климато-географические условия проживания монголов. Город Улан-Батор расположен на межгорном плато на высоте 1300–1350 м над уровнем моря, что на 600–800 м выше, чем Улан-Удэ [10].
Генотипическое разнообразие HCV было исследовано на основании анализа 200 ПЦР-позитивных проб крови больных ХГС, проживающих в Бурятии, и 200 — в Монголии (табл. 3). Установлено, что у монголов, больных ХГС, преобладает HCV-1 (98,0%), что значимо выше (р<0,001), чем у бурят (66,0%). Удельный вес других генотипов в каждой из сравниваемых групп был существенно меньше. Так, на НСV-2 и НСV-3 приходилось 2,0% у монголов и 34,0% — у бурят (р<0,001). Кроме указанных вариантов выявлены единичные пациенты с сочетанием двух и более генотипов, а также с нетипируемыми генотипами HCV, которые были исключены из статистического анализа с целью сохранения репрезентативности выборки.
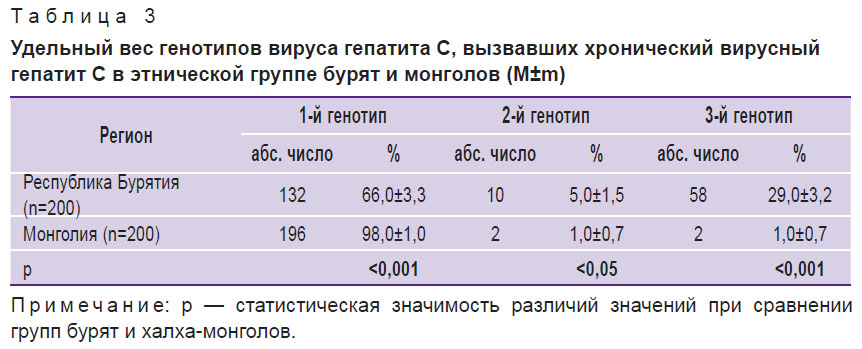
|
Таблица 3. Удельный вес генотипов вируса гепатита С, вызвавших хронический вирусный гепатит С в этнической группе бурят и монголов (M±m) |
На первом этапе работы проведено сравнение распространенности вариабельных сайтов изучаемых генов в популяциях здоровых лиц в этнических группах бурят и монголов (табл. 4).
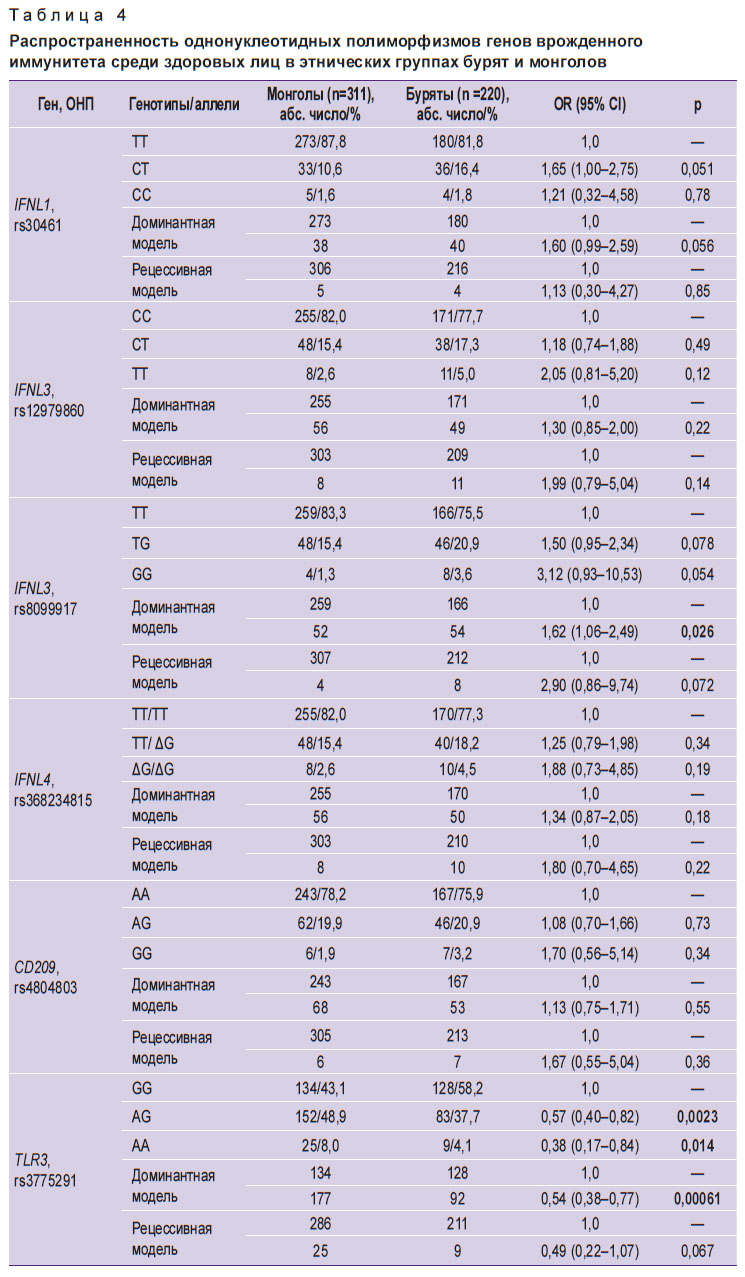
|
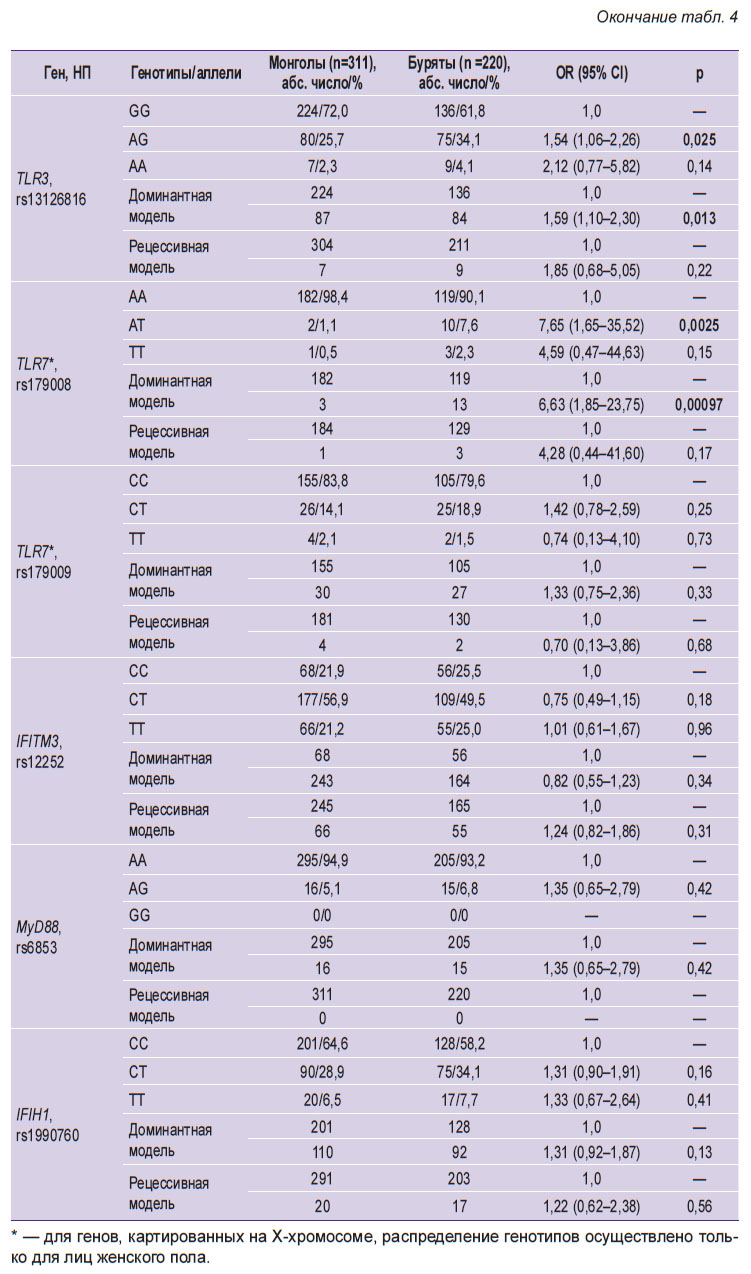
|
Таблица 4. Распространенность однонуклеотидных полиморфизмов генов врожденного иммунитета среди здоровых лиц в этнических группах бурят и монголов |
Установлены существенные различия в частоте встречаемости у этих лиц одного ОНП гена, кодирующего синтез интерферона-λ3, и трех ОНП генов, кодирующих синтез толл-подобных эндосомальных паттерн-распознающих рецепторов (TLR). Так, у здоровых монголов реже встречался GG-генотип TLR3 rs3775291 (OR=0,54; CI=0,38–0,77; p=0,0006). Вместе с тем значительно чаще встречались ТТ-генотип IFNL3 rs8099917 (OR=1,62; СI=1,06–2,49; p=0,026), GG-генотип TLR3 rs13126816 (OR=1,59; CI=1,10–2,30; p=0,013) и АА-генотип TLR7 rs179008 (OR=6,63; CI=1,85–23,75; p=0,00097). Последний из указанных генотипов отличался также тем, что 98,4% представителей этнической группы монголов являлись носителями доминантных АА-гомозиготных вариантов и лишь у трех обследованных в составе генотипа был обнаружен минорный Т-аллель.
Выявленные различия в частоте встречаемости этих четырех кандидатных генов у здоровых лиц обеих этнических групп послужили основанием для их дальнейшего изучения как потенциальных генетических факторов предрасположенности к инфицированию HCV-1 или толерантности к инфицированию HCV-2/3. Сравнительный анализ ОНП указанных генов в этнических группах монголов и бурят, больных ХГС, вызванным HCV-1, установил однотипные закономерности. В обеих группах значимо реже, чем у здоровых лиц, встречался ТТ-генотип IFNL3 rs8099917 (соответственно 65,9 и 77,7% у бурят; р=0,005 и 73,5 и 83,3% у монголов; р=0,002) и GG-генотип TLR3 rs3775291 (47,0 и 61,8% у бурят; р=0,007 и 56,1 и 72,0% у монголов; р=0,0002).
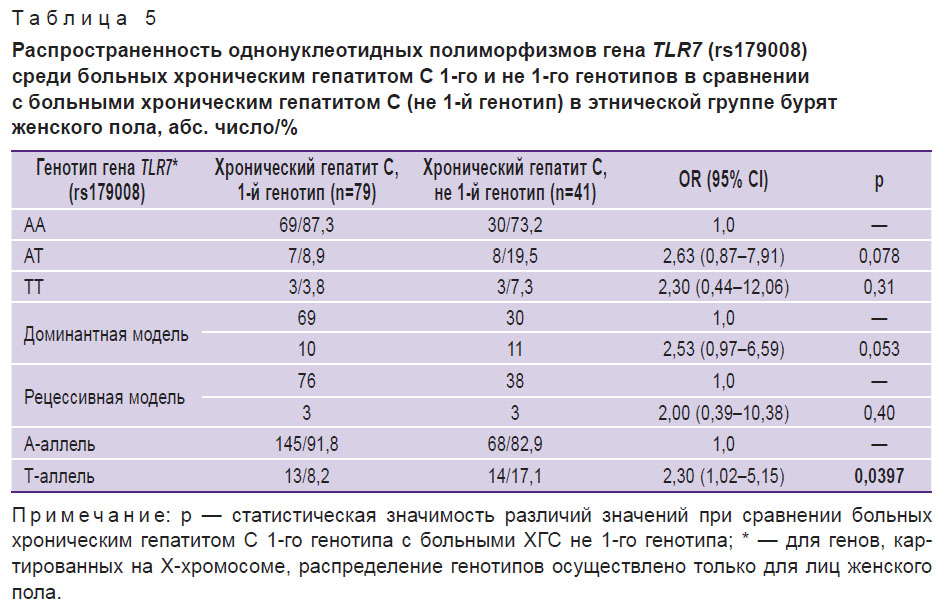
Сравнение генотипического разнообразия ОНП генов IFNL3 (rs8099917), TLR3 (rs3775291 и rs13126816), TLR7 (rs179008) у больных ХГС, инфицированных HCV-1 и HCV-2/3, было выполнено только в когорте больных бурят, так как в Монголии выявляются лишь единичные больные ХГС с не 1-м генотипом вируса. Проведенный анализ не позволил установить наличие ассоциации между генотипами вируса и генотипами ОНП изучаемых генов. Однако анализ по аллелям выявил значимо более высокую встречаемость Т-аллеля среди женщин-бурят, инфицированных не 1-м генотипом вируса (табл. 5).
Обсуждение
Исследование показало, что структура генотипов HCV существенно отличалась в Монголии и в соседней Бурятии. На территории Монголии обнаружено доминирующее преобладание HCV-1 (98,0%), что значимо выше (р<0,001) его распространенности на территории Бурятии (66,0%). В условиях полной сопоставимости групп по клинико-лабораторным и половозрастным показателям естественно предположить, что причиной доминирования 1-го генотипа вируса среди монголов может быть полиморфизм генов врожденного иммунитета, которые создают селективные преимущества для распространения данного генотипа или препятствуют циркуляции HCV-2/3 за счет генетической толерантности к инфицированию. Первыми работами в этом направлении, выполненными на этнических группах европеоидов, были исследования P. Wietzke-Braun с соавт. (2006) и E. Askar с соавт. (2009) [15, 16]. В одной из них был описан ОНП гена интерферон-регулирующего фактора IRF-1, который обеспечивает генетическую резистентность к HCV-3a [15]. В другой работе был обнаружен гомозиготный генотип ТТ rs3775291 гена TLR3, носители которого не болеют HCV-1a [16]. Поиски механизма этого явления привели авторов к выводу, что в основе резистентности некоторых людей к HCV-1a могут лежать активные реакции врожденного иммунитета. У лиц европеоидной расы, имеющих ТТ-генотип rs3775291, в ответ на индукторы обнаружена выраженная экспрессия гена TLR3 в клетках печени, обеспечивающая последующую активацию интерферон-регулирующего фактора-3 (IRF-3) и ядерного фактора κB(NF-κB), что приводит к интенсивной выработке интерферона-β и провоспалительных цитокинов [16].
В настоящей работе среди генетических факторов, которые могут оказывать влияние на формирование структуры циркулирующих генотипов HCV в популяции бурят и монголов, выявлены ОНП трех генов (IFNL3, TLR3 и TLR7), частота встречаемости которых существенно отличалась в изучаемых когортах.
Генотипы вариабельных сайтов IFNL3 относятся к одним из первых полиморфизмов, описанных при вирусном гепатите С, имеющих ассоциацию со спонтанным клиренсом вируса и ответом на противовирусное лечение препаратами интерферона и рибавирина [17]. В настоящем исследовании установлено, что благоприятный в отношении спонтанного клиренса HCV генотип ТТ rs8099917 IFNL3 существенно чаще встречается в популяции здоровых монголов, чем у бурят.
Другие ОНП, распределение которых значимо отличалось среди здоровых монголов и бурят, были локализованы в генах TLR3 (rs3775291, rs13126816) и TLR7 (rs179008). TRL относятся к паттерн-распознающим рецепторам и принимают активное участие в запуске и регуляции цитокинового воспаления. TLR3- и TLR7-рецепторы локализованы трансмембранно на эндосоме, куда попадает вирус после интернализации. Эти рецепторы распознают одно-, двуспиральную РНК, которая образуется в процессе репликации и сборки частиц практически всех РНК-содержащих вирусов. Полиморфизм генов TLR может оказывать влияние как на распознавание вируса (процессинг), так и на активацию сигнального пути, вызывающего воспаление (сигналинг). Сравнение полиморфизмов обоих генов показало существенные различия в частоте их встречаемости в сравниваемых этнических группах. Таким образом, обследование практически здорового населения монголов и бурят выявило три кандидатных гена, ОНП которых могут оказывать влияние на структуру генотипов HCV, циркулирующих в популяции.
Дальнейшие исследования были сосредоточены на анализе вышеуказанных четырех полиморфизмов в трех кандидатных генах IFNL3, TLR3 и TLR7 в популяции монголов и бурят, больных ХГС, вызванным различными генотипами вируса. Более частая встречаемость благоприятного генотипа ТТ rs8099917 гена IFNL3 среди здоровых лиц в сравнении с больными ХГС, вызванным HCV-1, как среди монголов, так и среди бурят, подтверждает хорошо известный факт генетической устойчивости носителей данного генотипа к инфицированию HCV [17]. Как показали полученные данные, исследование использования ОНП rs8099917 гена IFNL3 как прогностического фактора, применяемого для европеоидов и негроидов, приемлемо и для этнических групп монголоидов, проживающих на территории Бурятии и Монголии.
Кроме ОНП гена IFNL3 был обнаружен еще один вариабельный сайт rs13126816 гена TLR3, GG-генотип которого маркирует устойчивость к заболеванию вирусным гепатитом С, вызванным HCV-1. Этот ОНП также был универсальным и касался обеих этнических групп. Носители GG-генотипа имели меньший риск заболевания по сравнению с остальными лицами: среди монголов — в два раза, а среди бурят — в 1,8 раза. Предшествующее исследование этого ОНП у европеоидов [18] показало, что макрофаги людей, носителей G-аллеля, способны к быстрой и мощной продукции интерферона-β. По данным исследования, выполненного в США в этнических группах европеоидов, афроамериканцев и латиноамериканцев [18], G-аллель rs13126816 гена TLR3 был ассоциирован с высокой частотой спонтанного клиренса HCV-1. Этот же ОНП ассоциирован с устойчивостью индивидуумов к вирусу герпеса [19]. Таким образом, в исследуемых группах обнаружены два ОНП (rs8099917 IFNL3 и rs13126816 TLR3), дикие генотипы которых определяют устойчивость их носителей к инфицированию HCV-1.
Поиск генотипов генов, вовлеченных в реакции врожденного иммунитета, у больных ХГС, вызванным 1-м и 2/3-м генотипами HCV, не выявил их существенных отличий в группах. Вместе с тем анализ по отдельным аллелям установил, что Т-аллель rs179008 гена TLR7 более чем в два раза чаще встречается у женщин, больных ХГС, вызванным 2/3-м генотипом, чем 1-м (р=0,04). ОНП rs179008 гена TLR7 локализован в 3-м экзоне Х-хромосомы. По своему характеру данный ОНП является несинонимическим и характеризуется заменой в кодируемом белке глютамина (Gln) на лейцин (Leu). В результате сигнальный пептид рецептора TLR7 проявляет функциональную неполноценность [20]. В ответ на синтетические индукторы в дендритных клетках и гепатоцитах носителей минорного Т-аллеля наблюдается низкая экспрессия мРНК IFNL1, IL-10Rβ и IL-28Rα [21]. Кроме этого, Т-аллель ассоциирован с высокой вирусной нагрузкой, плохим ответом на противовирусную терапию и предрасположенностью к быстрому прогрессированию ВИЧ-инфекции у женщин [21, 22]. Мононуклеары здоровых людей, носителей Т-аллеля, in vitro слабо продуцируют IFN-α в ответ на индукторы интерферона [22]. Очевидно, более частая встречаемость мутантного гена TLR7 у женщин, больных ХГС, вызванным не 1-м генотипом вируса, является следствием слабого интерферонового и цитокинового ответа на инфицирование.
Как известно, HCV-2/3 проявляют более высокую чувствительность к IFN-α в отличие от HCV-1 и HCV-4. Генетическое нарушение синтеза IFN-α в ответ на инфицирование HCV-2/3 является весьма значимым фактором обеспечения длительной персистенции данных генотипов вируса. Редкое присутствие среди населения Монголии ТТ-генотипа rs179008 гена TLR7 (0,5%) определяет популяционную устойчивость населения к инфицированию HCV-2/3 и обусловливает абсолютное доминирование HCV-1. Причины отсутствия минорного мутантного Т-аллеля rs179008 гена TLR7 требуют дополнительных исследований и могут лежать у населения Монголии в основе особенностей структуры генотипов не только HCV, но и других вирусов, при которых реакции врожденного иммунитета играют существенную роль. Определенное значение может иметь и копийность гена TLR7, частота встречаемости которой среди населения Монголии остается неизученной.
Заключение
Проведенные исследования позволили обнаружить кандидатный ген TLR7, вариабельные сайты которого (rs179008) с различной частотой встречаются у женщин-бурят, больных 1-м и не 1-м (2/3) генотипами вируса. Т-аллель более чем в два раза чаще встречается у больных не 1-м генотипом вируса (17,1 и 8,2% соответственно). Низкая распространенность 2/3-го генотипов HCV на территории Монголии может быть обусловлена редкой встречаемостью среди населения мутантного Т-аллеля TLR7 (rs179008), ассоциированного с предрасположенностью к инфицированию HCV-2/3, что в настоящей работе продемонстрировано на примере этнической группы бурят.
Финансирование исследования. Работа выполнена за счет средств гранта Российского фонда фундаментальных исследований №16-54-44047.
Конфликт интересов. У авторов статьи конфликта интересов не имеется.
Литература
- Lavanchy D. Evolving epidemiology of hepatitis C virus. Clin Microbiol Infect 2011; 17(2): 107–115, https://doi.org/10.1111/j.1469-0691.2 010.03432.x
- Global hepatitis report 2017. Geneva: World Health Organization; 2017.
- Gower E., Estes C., Blach S., Razavi-Shearer K., Razavi H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J Hepatol 2014; 61(1 Suppl): S45–S57, https://doi.org/10.1016/j.jhep.2014.07.027.
- Baatarkhuu O., Kim D.Y., Ahn S.H., Nymadawa P., Dahgwahdorj Y., Shagdarsuren M., Park J.Y., Choi J.W., Oyunbileg J., Oyunsuren T., Han K.H. Prevalence and genotype distribution of hepatitis C virus among apparently healthy individuals in Mongolia: a population-based nationwide study. Liver Int 2008; 28(10): 1389–1395, https://doi.org/10.1111/j.1478-3231.2008.01820.x.
- Малов С.И. Сравнительная клинико-эпидемиологическая характеристика вирусного гепатита C на сопредельных территориях России и Монголии. Автореф. дис. … канд. мед. наук. М; 2017.
- Никитина Г.Ю., Семененко Т.А., Готвянская Т.П., Хахаева И.Б., Коноплева М.В., Николаева О.Г., Ярош Л.В., Кожевникова Л.К., Суслов А.П. Частота выявления маркеров инфицирования вирусами парентеральных гепатитов среди медицинских работников в регионах Российской Федерации с различной интенсивностью эпидемического процесса. Клиническая микробиология и антимикробная химиотерапия 2017; 19(2): 161–167.
- Ryan E.J., Dring M., Ryan C.M., McNulty C., Stevenson N.J., Lawless M.W., Crowe J., Nolan N., Hegarty J.E., O’Farrelly C. Variant in CD209 promoter is associated with severity of liver disease in chronic hepatitis C virus infection. Hum Immunol 2010; 71(8): 829–832, https://doi.org/10.1016/j.humimm.2010.05.007.
- Jiménez-Sousa M.A., Rallón N., Berenguer J., Pineda-Tenor D., López J.C., Soriano V., Guzmán-Fulgencio M., Cosín J., Retana D., García-Álvarez M., Miralles P., Benito J.M., Resino S. TLR3 polymorphisms are associated with virologic response to hepatitis C virus (HCV) treatment in HIV/HCV coinfected patients. J Clin Virol 2015; 65: 62–67, https://doi.org/10.1016/j.jcv.2015.02.004.
Barkhash A.V., Perelygin A.A., Babenko V.N., Brinton M.A., Voevoda M.I. Single nucleotide polymorphism in the promoter region of the CD209 gene is associated with human predisposition to severe forms of tick-borne encephalitis. Antiviral Res 2012; 93(1): 64–68, https://doi.org/10.1016/j.antiviral.2011.10.017.- Кручкин Ю.Н. Современная Монголия. Энциклопедический справочник. Улан-Батор; 2011; 1154 c.
- Симбирцев А.С. Цитокины в патогенезе инфекционных и неинфекционных заболеваний человека. Медицинский академический журнал 2013; 13(3): 18–41.
- Obaid A., Ahmad J., Naz A., Awan F.M., Paracha R.Z., Tareen S.H., Anjum S., Raza A., Baumbach J., Ali A. Modeling and analysis of innate immune responses induced by the host cells against hepatitis C virus infection. Integr Biol (Camb) 2015; 7(5): 544–559, https://doi.org/10.1039/c4ib00285g.
- Akaike H. A new look at the statistical model identification. IEEE Transactions on Automatic Control 1974; 19: 716–723, https://doi.org/10.1109/tac.1974.1100705.
- Певницкий Л.А. Статистическая оценка ассоциаций HLA-антигенов с заболеваниями. Вестник АМН СССР 1988; 7: 48–51.
- Wietzke-Braun P.,
Maouzi A.B., Mänhardt L.B., Bickeböller H., Ramadori G., Mihm S. Interferon regulatory factor-1 promoter polymorphism and the outcome of hepatitis C virus infection. Eur J Gastroenterol Hepatol 2006; 18(9): 991–997, https://doi.org/10.1097/01.meg.0000224478.89545.76. - Askar E., Bregadze R., Mertens J., Schweyer S., Rosenberger A., Ramadori G., Mihm S. TLR3 gene polymorphisms and liver disease manifestations in chronic hepatitis C. J Med Virol 2009; 81(7): 1204–1211, https://doi.org/10.1002/jmv.21491.
- Tanaka Y., Nishida N., Sugiyama M., Kurosaki M., Matsuura K., Sakamoto N., Nakagawa M., Korenaga M., Hino K., Hige S., Ito Y., Mita E., Tanaka E., Mochida S., Murawaki Y., Honda M., Sakai A., Hiasa Y., Nishiguchi S., Koike A., Sakaida I., Imamura M., Ito K., Yano K., Masaki N., Sugauchi F., Izumi N., Tokunaga K., Mizokami M. Genome-wide association of IL28B with response to pegylated interferon-alpha and ribavirin therapy for chronic hepatitis C. Nat Genet 2009; 41(10): 1105–1109, https://doi.org/10.1038/ng.449.
- Qian F., Bolen C.R., Jing C., Wang X., Zheng W., Zhao H., Fikrig E., Bruce R.D., Kleinstein S.H., Montgomery R.R. Impaired toll-like receptor 3-mediated immune responses from macrophages of patients chronically infected with hepatitis C virus. Clin Vaccine Immunol 2013; 20(2): 146–155, https://doi.org/10.1128/cvi.00530-12.
- Svensson A., Tunbäck P., Nordström I., Padyukov L., Eriksson K. Polymorphisms in toll-like receptor 3 confer natural resistance to human herpes simplex virus type 2 infection. J Gen Virol 2012; 93(Pt 8): 1717–1724, https://doi.org/10.1099/vir.0.042572-0.
- Fakhir F.Z., Lkhider M., Badre W., Alaoui R., Meurs E.F., Pineau P., Ezzikouri S., Benjelloun S. Genetic variations in toll-like receptors 7 and 8 modulate natural hepatitis C outcomes and liver disease progression. Liver Int 2018; 38(3): 432–442, https://doi.org/10.1111/liv.13533.
- Askar E., Ramadori G., Mihm S. Toll-like receptor 7 rs179008/Gln11Leu gene variants in chronic hepatitis C virus infection. J Med Virol 2010; 82(11): 1859–1868, https://doi.org/10.1002/jmv.21893.
- Schott E., Witt H., Neumann K., Bergk A., Halangk J., Weich V., Müller T., Puhl G., Wiedenmann B., Berg T. Association of TLR7 single nucleotide polymorphisms with chronic HCV-infection and response to interferon-a-based therapy. J Viral Hepat 2008; 15(1): 71–78, https://doi.org/10.1111/j.1365-2893.2007.00898.x.








