Дисфункция автономной нервной системы в патогенезе септических критических состояний (обзор)
Возникающая дисфункция автономной нервной системы (АНС) головного мозга при сепсисе обусловливает летальность и тяжесть системного воспаления. Известны многочисленные данные о влиянии АНС на возникновение, течение и исход генерализованной инфекции. Парасимпатическое звено АНС моделирует воспаление через холинорецепторы внутренних органов, макрофагов и лимфоцитов (холинергический противовоспалительный путь — cholinergic anti-inflammatory pathway). Симпатическое звено АНС также изменяет активность макрофагов и лимфоцитов путем влияния на β2-адренорецепторы, вызывая экспрессию внутриклеточных генов, ответственных за синтез цитокинов (beta2-adrenergic receptor interleukin-10 anti-inflammatory pathway, β2AR–IL-10). Взаимодействие АНС, инфекционных агентов и иммунной системы обеспечивает поддержание гомеостаза или появление критической генерализованной инфекции. Во время воспаления активация АНС моделирует воспалительный ответ через освобождение симпатических или парасимпатических нейротрансмиттеров и нейропептидов. Чрезвычайно важно определять функциональное состояние АНС при критических состояниях, так как в роли противо- или провоспалительных стимулов могут выступать как холинергические, так и симпатомиметические агенты.
Введение
Функциональное состояние автономной нервной системы (АНС) играет важную роль в регуляции воспалительного ответа организма [1]. С учетом этого патогенность возбудителя, его вид, путь инвазии нельзя считать главными факторами возникновения и поддержания генерализованной инфекции. Именно нейроэндокринный и иммунный ответы организма на бактериальную инвазию оказывают определяющее влияние на развитие и прогноз септического состояния, полиорганную дисфункцию и летальность.
Патофизиологические аспекты влияния функционального состояния автономной нервной системы на течение сепсиса и септического шока
Дисфункция АНС — триггер патофизиологии и клиники сепсиса. При септических состояниях системный воспалительный синдром (systemic inflammatory response syndrome, SIRS) и мультиорганный дисфункциональный синдром (multiorgan dysfunction syndrome, MODS) наиболее тяжело проявляются именно при имбалансе АНС [2]. Дисфункция АНС служит главным триггером патофизиологических расстройств и клиники сепсиса [3–5]. В функциональном статусе АНС выделяют два регуляторных компонента: вагусный (парасимпатическое звено) и симпатоадреналовый (симпатическое звено). Многочисленные данные свидетельствуют, что усиление парасимпатического компонента обеспечивает снижение воспалительных проявлений через реализацию так называемого холинергического антивоспалительного пути (cholinergic anti-inflammatory pathway, CAP). В ряде научных публикаций раскрыты механизмы влияния CAP на системные и локальные воспалительные реакции [6, 7]. При развитии воспаления афферентная импульсация передается по блуждающему нерву от периферии к стволовым структурам головного мозга. Вагусная эфферентная передача переносит исходящие сигналы, которые заканчиваются в селезенке, печени, кишечнике и других органах [8–16]. Стимуляция эфферентного блуждающего нерва, как и медикаментозная холинергическая терапия (применение блокаторов холинэстеразы) приводит к выделению ацетилхолина в эти органы. В свою очередь ацетилхолин взаимодействует с никотиновыми ацетилхолиновыми рецепторами (nAChR), в частности с никотиновым ацетилхолиновым рецептором α7 (α7nAChR), который экспрессируется макрофагами и другими клетками, продуцирующими цитокины. Ацетилхолин дозозависимо понижает продукцию TNF-α макрофагами человека, и этот антивоспалительный эффект реализуется через α7-субъединицу никотинового холинорецептора. Стимуляция ацетилхолином α7nAChR оказывает ингибирующее действие на лейкоциты, включая макрофаги и врожденные лимфоидные клетки 2-го типа. Врожденные лимфоидные клетки — это группа лимфоцитов, которые вовлечены в быстрое цитокинзависимое реагирование организма во время воспалительного процесса и, в отличие от обычных лимфоцитов приобретенного иммунитета, у них отсутствуют антигенспецифические рецепторы, они могут реагировать на широкий спектр воспалительных стимулов. В конечном счете концентрация провоспалительных цитокинов заметно ингибируется. Это нейроиммунное сообщение и есть холинергический антивоспалительный путь [17].
Важно отметить, что и симпатическое звено АНС также активно регулирует воспалительные реакции. Открыт механизм нейрогенного торможения воспаления, который основан на продукции норэпинефрина катехоламинергическими волокнами селезенки. Норэпинефрин связывается с β2-адренергическими рецепторами (β2AR) CD4+ T-клеток(Т-хелперов). CD4+ запускают высвобождение ацетилхолина, ингибирующего секрецию воспалительных цитокинов макрофагами по известному пути — посредством передачи сигналов α7nAChR с последующей реализацией CAP. Кроме того, агонисты β2AR вызывают увеличение продукции интерлейкина IL-10 миелоидными клетками с противовоспалительными свойствам — β2AR-противовоспалительный путь (beta-2 adrenergic receptor interleukin-10 anti-inflammatory pathway, β2AR–IL-10) [18]. На примере активации макрофагов при персистировании диабета показана роль β2-агонистов (медиаторов симпатической нервной системы) в блокировании системных воспалительных реакций, что в свою очередь приводит к торможению развития макроангиопатий [19]. Агонисты β2AR выступают как мощные ингибиторы продукции TNF-α макрофагами костного мозга. Противовоспалительное действие β2AR представлено на модели острого нарушения мозгового кровообращения. Ишемический инсульт провоцирует нейровоспалительный процесс и постоянное освобождение эпинефрина и норэпинефрина симпатической нервной системой. Повышенная β2-адренергическая передача сигналов после начала инсульта обычно подавляет реакцию микроглии, уменьшая активацию как провоспалительных, так и противовоспалительных цитокинов. Напротив, снижение β2-адренергической передачи сигналов в микроглии увеличивает как провоспалительную, так и противовоспалительную экспрессию цитокинов после инсульта. Следовательно, β2AR может представлять собой терапевтическую мишень для улучшения процесса протекания инсульта и воспаления после него [20].
Симпатические и парасимпатические противовоспалительные действия реализуются как минимум через активацию трех внутриклеточных сигнальных путей. Регуляция внутриклеточных сигнальных путей в лимфоцитах является критической для клеточного гомеостаза и иммунного ответа. Доказано, что β2AR ингибируют продукцию NF-κB (nuclear factor κB) — транскрипционного фактора, вызывающего экспрессию генов, ответственных за реализацию синтеза TNF-α, IL-1, и тем самым снижают воспалительный ответ (первый внутриклеточный механизм) [21]. Второй внутриклеточный механизм — это внутриклеточная сигнальная система JAK/STAT (janus kinases — signal transducer and activator of transcription) — менеджер последующей цитокиновой продукции, которая также активно моделирует воспалительный ответ. JAK/STAT-система представляет собой сигнальный путь, состоящий из янус-киназы, сигнального белка-трансдуктора и активатора транскрипции. Этот путь передает информацию от внеклеточных полипептидных сигналов через трансмембранные рецепторы непосредственно к промоторам генов-мишеней в ядре, где они связываются с соответствующими регуляторными последовательностями генов и запускают их транскрипцию [22]. Наконец, третий внутриклеточный сигнальный путь — PI3K/AKT/mTOR, центральными компонентами которого являются ферменты фосфоинозитид-3-киназа (рhosphoinositide 3-kinases,PI3K), киназы AKT (alpha serine/threonine-protein kinases) и mTOR (mammalian target of rapamycin), — мишень рапамицина млекопитающих. Представлены доказательства связи между рецептором IL-2 и β2AR. Обработка линий лимфоидных клеток человека одним только агонистом β2AR изопротеренола (ISO) увеличивает уровни цАМФ и опосредует стимулирующий ответ путем активации AKT и ERK, что повышает жизнеспособность клеток. Посредством этого молекулярного механизма передача сигналов β2AR может как стимулировать, так и подавлять ответы лимфоцитов и таким образом объяснять, как определенные терапевтические агенты могут влиять на иммунную реакцию [23] (рис. 1).
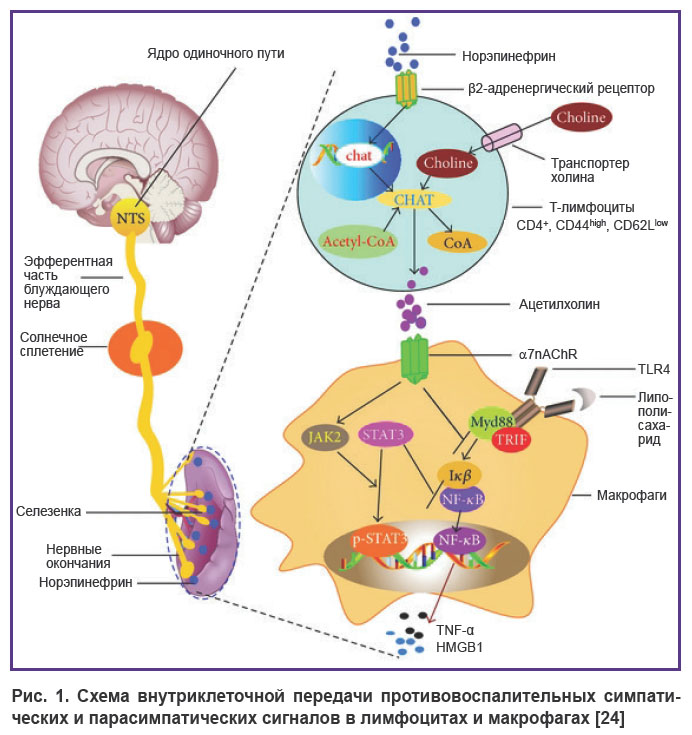
|
Рис. 1. Схема внутриклеточной передачи противовоспалительных симпатических и парасимпатических сигналов в лимфоцитах и макрофагах [24] |
Взаимосвязь автономной нервной системы и микробиоты кишечника. Их взаимодействие чрезвычайно важно в процессах развития септических состояний. A.C. Vaughn с соавт. [25] показали, чтоэнергоемкая диета вызывает изменения микробиоты кишечника и посредством парасимпатической афферентной импульсации нарушает взаимодействие мозга и кишечника, приводя к накоплению жира. Около 100 000 млрд. бактерий находится в кишечнике человека. Композиция бактериоты зависит от возраста, массы тела, диеты. В норме бактериота защищает организм от пенетрации патогенных микроорганизмов и выполняет следующие функции: отвечает за интеграцию функции кишечной стенки, чувствительность к инсулину, метаболизм и, как установлено в последнее время, обусловливает взаимодействие со структурами головного мозга [26]. Липополисахариды из грамотрицательных бактерий проникают через стенку кишечника в кровь. Этот процесс индуцирует эндотоксемию, воспаление, нарушает метаболизм глюкозы, вследствие чего появляются резистентность к инсулину, ожирение, метаболический синдром, диабет 2-го типа, воспаление стенки кишечника, аутоиммунные процессы и карциногенез. Норадреналин, освобождаемый из терминальных синапсов АНС, повреждает продукцию протеина в слепой кишке. Активация стрессом гипоталамо-гипофизарно-надпочечниковой (ГГН) оси и АНС увеличивает содержание кортизола и провоспалительных цитокинов, таких как TNF-α, IL-8, IL-1β, IL-6 [27–29]. Увеличение микробиологической нагрузки в кишечнике, избыточное освобождение цитокинов нарушают взаимодействие между структурами системы ГГН, АНС и кишечником (рис. 2).
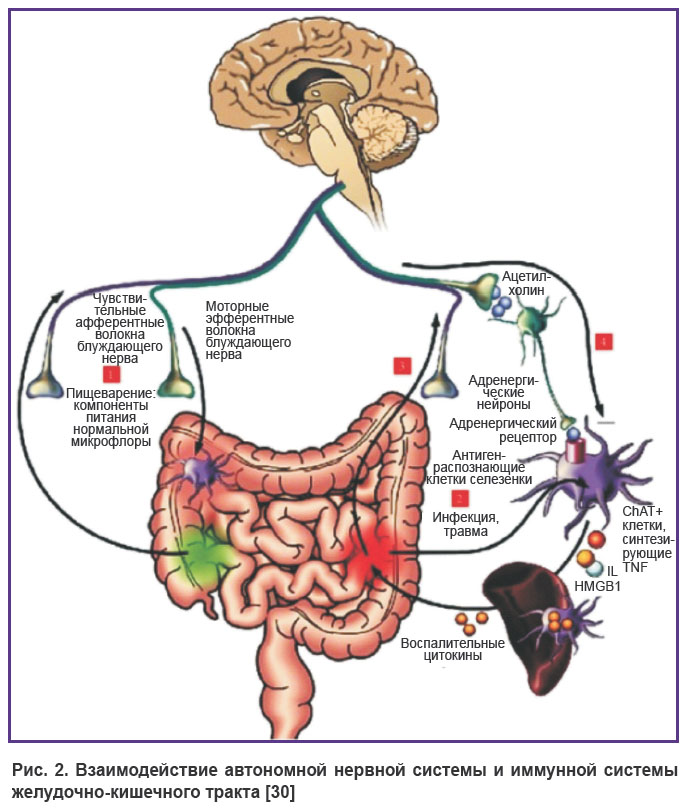
|
Рис. 2. Взаимодействие автономной нервной системы и иммунной системы желудочно-кишечного тракта [30] |
Анализ вариабельности ритма сердца — «золотой стандарт» оценки функционального состояния автономной нервной системы. В последних научных публикациях, посвященных анализу симпатовагусных влияний при сепсисе, большое место занимает неинвазивный анализ функционального статуса АНС путем изучения вариабельности ритма сердца (ВРС). Проведенный метаанализ с оценкой 2283 наблюдений показал негативную взаимосвязь параметров временнόго анализа ВРС (SDNN), высокочастотного спектра ВРС и воспалительных маркеров [31]. Получены данные, что усиление парасимпатического компонента (более высокая ВРС) обеспечивает понижение воспаления через реализацию CAP [32–34].
Важно иметь в виду еще один аспект при развитии системного воспаления и появления SIRS и MODS. Изучение IL-1β, IL-6, TNF-α обусловлено задачей оценки развития так называемого антивоспалительного синдрома (compensatory anti-inflammatory response syndrome, CARS), что препятствует развитию SIRS [35]. Возникновение и прогрессирование сепсиса вызвано дисрегуляторным ответом организма «хозяина» на инфекцию, что обусловливает органную дисфункцию и летальность. Во время сепсиса тканевое повреждение приводит к контролируемой активации комплемента, коагуляционным расстройствам, дисфункции тромбоцитов, гиперпродукции цитокинов. Баланс между SIRS и CARS определяет исход сепсиса [36]. Удержание сбалансированного ответа АНС в этих условиях — основа формирования компенсаторного антивоспалительного синдрома [37, 38]. Важен контроль (в том числе на основе цифрового навигационного подхода к параметрам нормы и патологии ВРС) функции АНС для лечения и профилактики МОDS и SIRS.
Органная дисфункция и патология автономной нервной системы при сепсисе
Автономная нервная система при патологии головного мозга. В структурах головного мозга (гипоталамус, гиппокамп, nucleus tractus solitarius, гипофиз) находятся основные центры АНС, которые в первую очередь повреждаются при развитии различных патологических состояний этого анатомического региона (черепно-мозговой травмы, нарушения мозгового кровообращения, аноксии и т.д.). Мозг — мишень для медиаторов системной воспалительной реакции и активных форм кислорода. К настоящему времени описаны аутоиммунная форма эпилепсии, аутоиммунный анти-NMDA рецепторный энцефалит, аутоиммунные антитела к холинорецепторам соматической нервной системы и АНС [39–41]. Вместе с тем клетки микроглии являются макрофагами костномозгового кроветворения, а клетки астроглии способны синтезировать активные формы кислорода и TNF-α. Микроглия вызывает каскад реакций, приводящих к вторичному поражению ЦНС. S.S. Gaddam и соавт. [42] отмечали, что повреждения и дисфункция головного мозга после черепно-мозговой травмы вызывают умеренные или выраженные органные нарушения в других анатомических областях или системную инфекцию. M.G. Frasch с соавт. [43] в эксперименте оценили корреляцию некоторых параметров ВРС и уровней провоспалительных цитокинов при активации микроглии эмбрионального мозга в условиях гипоксии. Они установили, что среднеквадратичное различие между длительностью соседних R–R-интервалов (RMSSD) коррелирует с уровнем IL-1β в плазме (коэффициент корреляции положительный, r=0,57; p=0,02; n=7), с белками HMGB1 (nuclear high mobility group box 1 proteins) таламической микроглии (коэффициент корреляции отрицательный, r=–0,94; p=0,005) и с уровнем микроглиальных холинорецепторов макрофагов α7nAChR белого вещества головного мозга (коэффициент корреляции положительный, r=0,83; p=0,04). Данные результаты свидетельствуют о возможности оценки уровня нейровоспаления плода путем регистрации ВРС и о появляющихся возможностях для неинвазивного мониторинга и целенаправленного лечения. Большой интерес вызывают приведенные в работе A.J. Nicholls с соавт. [44] результаты влияния норадреналина на функциональную активность нейтрофилов через соединение с адренорецепторами. Нейтрофилы были изолированы из костного мозга в эксперименте на мышах, которые получали норадреналин в разных концентрациях. Воспроизводилась модель инсульта и оценивалась активность нейтрофилов (способность мигрировать под воздействием различных цитокинов) при применении норадреналина в течение 4 и 24 ч. Лечение норадреналином в течение 4 ч достоверно снижало хемотаксис нейтрофилов, вызывало супрессию продукции интерферона (IFN-γ) и IL-10, снижало активацию нейтрофилов и фагоцитоз. Эти данные показывают важность оценки функционального состояния АНС, где норадреналин может выступать как блокатором, так и индуктором системного воспаления [45].
Воспалительные заболевания кишечника негативно влияют на качество жизни миллионов пациентов во всем мире. Точная этиология заболевания остается неясной, основной причиной считается аберрантная активация иммунной системы. Наиболее перспективными являются методы лечения воспалительных заболеваний кишечника, которые избирательно ингибируют активацию иммунных клеток без широкой иммуносупрессии. Одним из таких способов лечения стало ингибирование активации иммунных клеток, предотвращающее выработку провоспалительных цитокинов посредством нервной стимуляции. Новые терапевтические подходы основаны на открытии CAP — рефлекторной дуги, которая индуцирует эфферентную передачу сигналов блуждающего нерва для снижения активации иммунных клеток и, следовательно, смертности во время сепсиса и септического шока [46, 47]. Активация CAP стимуляцией блуждающего нерва оказывает защитное действие при широком разнообразии клинических расстройств, включая болезнь Крона. Канонический путь CAP включает активацию α7nAChR — позитивных макрофагов селезенки — с помощью положительных β2AR CD4+ T-клеток. Ультразвуковая стимуляция или стимуляция блуждающего нерва также активировали α7nAChR-позитивные перитонеальные макрофаги, и адаптивный перенос этих активированных макрофагов снижал проявления колита [48].
Ревматоидный артрит. Это сложное хроническое мультисистемное аутоиммунное заболевание, при котором воспалительный каскад вызывает спазм сосудов и нарушение остеокластогенеза, что в конечном итоге приводит к разрушению костей и хрящей. В исследовании [49] оценена экспрессия и локализация гена α7nAChR в сердце, печени, селезенке, легких, почках и суставах в экспериментальной модели индуцированного артрита у крыс. При активации CAP-пути уровни экспрессии мРНК снижались, что приводило к уменьшению воспаления при данном виде патологии.
При хронической обструктивной болезни легких (ХОБЛ) активность холинергической системы повышается, что служит одной из причин ограничения воздушного потока, вызванного сокращением гладких мышц дыхательных путей. Следовательно, блокирование сократительных действий антихолинергическими средствами является полезным терапевтическим вмешательством для уменьшения ограничения воздушного потока. В дополнение к известным эффектам бронхоконстрикции и секреции слизи накопленные данные, полученные на животных моделях ХОБЛ, позволяют предположить, что ацетилхолин играет роль в воспалении. В последнее время для лечения ХОБЛ стали доступны комбинации β2-адренергических агонистов длительного действия (long-acting β adrenoceptor agonists, LABA) и мускариновых антагонистов длительного действия (long acting muscarinic antagonists, LAMA). Эти двойные бронходилататоры длительного действия могут оказывать синергетическое противовоспалительное действие, поскольку стимуляция β2AR индуцирует ингибирующие эффекты в воспалительных клетках [50]. В дополнение к эффектам расслабления дыхательных путей также обнаружено, что агонисты β2AR обладают широкими противовоспалительными эффектами [51].
Бронхиальная астма характеризуется увеличением воспалительных клеток, особенно эозинофилов, а также продукцией активных форм кислорода, обусловленной высвобождением цитокинов, ассоциированных с Т-хелперами. САР ингибирует выработку цитокинов и контролирует воспаление. На модели аллергической астмы исследовано влияние фармакологической активации CAP неостигмином на окислительный стресс и воспаление дыхательных путей. Активация CAP неостигмином снижает уровни провоспалительных цитокинов (IL-4, IL-5, IL-13, IL-1β и TNF-α), что приводит к снижению притока эозинофилов в слизистой оболочке бронхов [52].
При воспалительных процессах в почках и печени CAP также играет терапевтическую роль. Важно отметить, что иммунные клетки в селезенке экспрессируют большинство компонентов холинергической системы, таких как ацетилхолин, холинацетилтрансфераза, ацетилхолинэстераза, а также мускариновые и никотиновые рецепторы ацетилхолина, что делает возможным взаимодействие между обеими системами. В целом эта коммуникация подавляет воспаление, достигаемое посредством различных механизмов в зависимости от вовлеченных клеток [53, 54].
В акушерской патологии на примере преэклампсии показано [55], что CAP регулирует системное воспаление посредством активации α7nAChR, экспрессируемых в моноцитах и макрофагах периферической крови. Моноциты периферической крови были выделены у 30 небеременных женщин (NP), 32 нормотензивных беременных (NT) и у 35 женщин с преэклампсией (PE). Обнаружено, что уровни экспрессии белка и мРНК α7nAChR в моноцитах у женщин с преэклампсией были значительно ниже, чем у женщин из групп NP и NT (в обоих случаях p<0,01). Уровни экспрессии белка α7nAChR в моноцитах отрицательно коррелировали с уровнями систолического артериального давления (r=–0,40; р=0,04), протеинурии (r=–0,54; р<0,01), TNF-α (r =–0,42; р=0,01) и IL-1β (r=–0,56; р<0,01) и положительно коррелировали с уровнями IL-10 (r=0,43; р=0,01) у женщин из группы PE. Повышение уровней TNF-α, IL-1β и IL-6 было выше в группе PE, чем в группах NP и NT (все p<0,01), но уровень IL-10 в группе PE был ниже, чем в группах NP и NT (p<0,01). Кроме того, активность NF-κB в моноцитах у женщин из группы PE была выше, чем у женщин из групп NP и NT (p<0,01). Эти результаты предполагают, что подавление α7nAChR может быть связано с развитием преэклампсии посредством увеличения провоспалительных и уменьшения высвобождения противовоспалительных цитокинов через путь NF-κB.
Диабет 1-го типа — это аутоиммунное заболевание, вызванное Т-клетками, которое приводит к гибели β-клеток поджелудочной железы и, следовательно, к потере выработки инсулина. На модели экспериментального аутоиммунного заболевания авторы работы [56] показали, что предварительная обработка специфическим ингибитором ацетилхолинэстеразы (AChEI) параоксоном предотвращала развитие гипергликемии у мышей. Это коррелировало с ингибированием инфильтрации Т-клеток в островки поджелудочной железы и снижением провоспалительных цитокинов. Холинергическая стимуляция обладает лечебным эффектом при аутоимунном диабете.
Ожирение является хроническим состоянием, связанным с дисфункцией АНС и ГГН-оси и умеренными воспалительными проявлениями. Моноциты у животных с ожирением, вызванным диетой с высоким содержанием жиров, имели более высокие уровни экспрессии всех провоспалительных цитокинов и более высокий процент моноцитов с провоспалительным фенотипом, чем у животных с низкокалорийной диетой. При этом β2-адренергическая стимуляция в моноцитах была противовоспалительной только у животных с ожирением, у которых на начальном этапе отмечалось провоспалительное состояние [57, 58].
Терапевтические интервенции при дисфункции автономной нервной системы и генерализованной инфекции
Холинергический противовоспалительный путьконтролируется через вагус и предупреждает повреждения клеток и тканей при гиперпродукции цитокинов. Активность парасимпатической нервной системы (вагусная активность) может быть скомпрометирована, что приводит к ослаблению CAP. Нормализация парасимпатической нервной системы представляется новой перспективной терапевтической целью — добиться подавления системных воспалительных изменений и тем самым улучшить прогноз у пациентов с MODS и SIRS [59]. Новым направлением интенсивной терапии следует признать применение лекарственных препаратов, позволяющих ликвидировать имбаланс АНС [60, 61]. Изменение продукции провоспалительных цитокинов при действии блокаторов холинэстеразы (галантамина) или ваготомии снижает в конечном итоге степень выраженности эндотоксемии и смертность при сепсисе [62].
Фармакологическая модуляция с применением галантамина как метода холинергической стимуляции показала эффективность в ослаблении воспаления, связанного с ожирением, нейровоспалением и метаболическими нарушениями [63].В исследовании I. Njoku и соавт. [64] показано, что галантамин, являясь ингибитором холинэстеразы и позитивным аллостерическим модулятором nAChRs, снижает когнитивный дефицит после черепно-мозговых травм.
Активация CAP проявляется и при использовании другого блокатора холинэстеразы — физостигмина [65]. Авторы изучили 20 пациентов с периоперационным септическим шоком вследствие интраабдоминальной инфекции. Группа «физостигмин» получала начальную дозу 0,04 мг/кг салицилата физостигмина с последующей непрерывной инфузией 1 мг/ч в течение 120 ч (5 дней); группу «плацебо» лечили 0,9% хлоридом натрия. Оценивали средний балл недостаточности органов (SOFA) во время лечения и до 14 дней после лечения.Средние значения SOFA составили 8,9±2,5 и 11,3±3,6 (среднее ± SD) для групп «физостигмин» и «плацебо» соответственно. С учетом возраста больных разница между средними значениями не была статистически значимой (–2,37; 95% ДИ: от –5,43 до 0,70; р=0,121). Требуемые дозы норэпинефрина в группе «физостигмин» оказались ниже (р=0,064), а более быстрое снижение частоты сердечных сокращений указывало на меньшую гемодинамическую нестабильность.
Установлена роль дексмедетомидина, α2-адренергического агониста, вызывающего симпатическую ингибицию и активацию CAP. Дексмедетомидин предотвращает апоптоз нейронов и ингибирует воспаление в данных ситуациях. Применение дексмедетомидина в эксперименте уменьшает уровень белка S100β, нейронспецифической енолазы, IL-6 в плазме крови и веществе головного мозга [66]. В экспериментальной работе [67] методом иммуногистохимии изучена экспрессия α7nAChR и IL-1β, TNF-α, белка S100β и мозгового нейротрофического фактора (brain-derived neurotrophic factor, BDNF) при использовании дексмедетомидина. Этот α2-адренергический агонист снижает экспрессию в гиппокампе уровня α7nAChR и IL-1β, TNF-α и S100β и повышает уровень BDNF. Системное воспаление у крыс, вызванное интраперитонеальной инъекцией 5,0 мг/кг липополисахарида (lipopolysaccharide), приводило к дефициту сознания, развитию нейровоспаления в гиппокампе. Дексмедетомидин при этом предотвращал индуцированную системным воспалением активацию микроглии [68]. Этот α2-адренергический агонист регулирует экспрессию генов, активацию клеточных каналов, освобождение трансмиттеров, процессы воспаления и клеточного апоптоза [69–75].
В последних исследованиях [76, 77] показано, что сепсис-индуцируемая сердечная дисфункция обусловлена в ряде моментов и симпатической гиперактивностью (sympathetic nerve overstimulation). В одноцентровом рандомизированном исследовании показано, что применение инфузии эсмолола (селективного β1-адреноблокатора) у пациентов с септическим шоком уменьшает летальность в течение 1 мес после операции. Блокада β-рецепторов при сепсисе снижает продукцию провоспалительных цитокинов, супрессирует гиперметаболический статус, поддерживает баланс глюкозы и улучшает проявления коагулопатии. Появилось достаточное количество исследований, подчеркивающих важнейшую роль гипоталамических структур, регулирующих периферическую иммунную систему и воспаление [78–84].
Ацетилхолин является ключевым противовоспалительным трансмиттером холинергического противовоспалительного пути. Проведены исследования, в которых оценена взаимосвязь уровня концентрации ацетилхолина в плазме крови и проявлений воспалительных реакций [85]. В исследовании G. Tao и соавт. [86] 113 пациентов были включены в проспективное исследование. Всем им был выполнен протокол раннего энтерального питания через 24–48 ч после поступления в отделение интенсивной терапии. В плазме крови исходно, на 1, 3, 5, 7-й дни проведения раннего энтерального питания изучены уровни ацетилхолина и маркеры воспаления TNF-α, IL-1β и IL-6. 28-дневная летальность составила 28,3% (32/113). По сравнению с исходным (15,6±2,8 нмоль/л) уровень ацетилхолина в плазме значительно повышался на 3-и сутки — до 18,6±6,7 нмоль/л, на 5-е сутки — до 19,3±6,2 нмоль/л и на 7-е сутки — до 19,7±4,3 нмоль/л (р<0,001). По сравнению с исходным (176,2±50,4 пг/мл) уровень TNF-α в плазме значительно снижался на 3-и сутки — до 144,0±77,4 пг/мл, на 5-е сутки — 127,3±51,8 пг/мл и на 7-е сутки — до 111,4±42,5 пг/мл (р<0,05). По сравнению с исходным уровень IL-1 в плазме значительно снизился к 7-м суткам (р<0,05), а IL-6 — на 5-е и 7-е сутки (р<0,05). Повышенные уровни ацетилхолина в плазме крови коррелируют с благоприятным прогнозом критического состояния.
Симпатомиметические трансмиттеры также активно моделируют воспалительные реакции. Показано, что норадреналин и адреналин дозозависимо супрессируют освобождение IL-27 из активированных макрофагов, что улучшает выживаемость при септическом шоке [87]. Полученные данные подтверждают важную роль АНС при остром воспалении. Лимфоидные органы богато иннервированы симпатической нервной системой, где происходит секреция трансмиттера норадреналина — β2-специфического агониста (β2-specific agonist). Иммунные клетки содержат адренергические рецепторы, позволяя симпатической нервной системе напрямую контролировать иммунную функцию. Норадреналин может ингибировать продукцию провоспалительного цитокина TNF-α и увеличивать продукцию противовоспалительного цитокина IL-10 из иммунных клеток как ответ на появление в крови липополисахарида, вызывающего эндотоксемию. Таким образом, норадреналин влияет на системное воспаление при сепсисе [88]. Симпатомиметик (фенилэфрин) может ингибировать сепсис-индуцированную сердечную дисфункцию, воспаление, повреждение митохондрий через активацию внутриклеточного сигнального пути PI3K/AKT/mTOR. В эксперименте перитонит и сепсис вызывали путем перевязки слепой кишки и ее пункцией, при этом применение фенилэфрина приводило к снижению продукции TNF-α, IL-6 и летальности [89]. Эти и другие данные демонстрируют, что проявления системного воспаления наиболее выражены при холинергической или адренергической дисфункции.
Полученные на сегодняшний день результаты показывают новые возможные подходы к целенаправленной терапии дисфункции АНС при сепсисе и других состояниях иммунной дисрегуляции [90–93]. Чрезвычайно важно определять вид дисфункции АНС — в виде симпатической или парасимпатической гиперактивности — для последующей целенаправленной таргет-терапии системного воспаления симпатомиметиками или симпатолитиками [94–98].
Заключение
Возникающая дисфункция автономной нервной системы головного мозга при сепсисе во многом обусловливает летальность и тяжесть системного воспаления [99–104]. Однако до сих пор нет специфических подходов к оценке дисфункции этой структуры головного мозга для выработки целевых (таргетных) подходов к ее коррекции. Приведенные данные со всей очевидностью показывают, что автономная нервная система влияет на возникновение, течение и исход генерализованной инфекции. Парасимпатическое звено автономной нервной системы моделирует воспаление через холинорецепторы внутренних органов, макрофагов и лимфоцитов (холинергический противовоспалительный путь — cholinergic anti-inflammatory pathway). Симпатическое звено автономной нервной системы также изменяет активность макрофагов и лимфоцитов путем влияния на β2-адренорецепторы, вызывая экспрессию внутриклеточных генов, ответственных за синтез цитокинов β2AR–IL-10. Таким образом, взаимодействие автономной нервной системы, инфекционных агентов и иммунной системы обеспечивает поддержание гомеостаза или появление критической генерализованной инфекции (рис. 3).
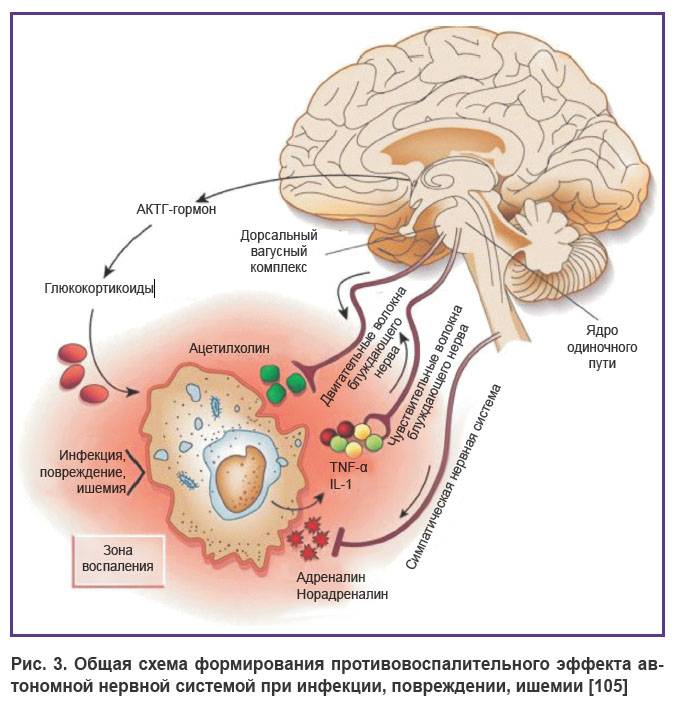
|
Рис. 3. Общая схема формирования противовоспалительного эффекта автономной нервной системой при инфекции, повреждении, ишемии [105] |
Во время воспаления активация автономной нервной системы моделирует воспалительный ответ через освобождение симпатических или парасимпатических нейротрансмиттеров и нейропептидов. Чрезвычайно важно определять функциональное состояние автономной нервной системы при критических состояниях, так как в роли противовоспалительных или провоспалительных стимулов могут выступать как холинергические, так и симпатомиметические агенты. Удержание сбалансированного ответа автономной нервной системы в этих условиях — основа формирования компенсаторного антивоспалительного синдрома (CARS), что препятствует развитию системного воспалительного синдрома SIRS. Моделируя активность симпатической и парасимпатической нервной системы через блокаду или активацию адрено- и холинорецепторов, можно добиться лечения воспаления при сепсисе. Задача современных научных исследований — определить вид дисфункции АНС и подойти к применению интенсивной терапии в целенаправленном таргетированном направлении. Очевидно, что активация этого холинергического противовоспалительного пути, ингибирующая продукцию TNF-α и других цитокинов, а также защищающая от воспалительных повреждений во время эндотоксемии и острого сепсиса, наиболее целесообразна в случае чрезмерной симпатической гиперактивности. В свою очередь активация β2AR-агонистами адренорецепторов наиболее целесообразна при парасимпатической гиперактивности. Изучение взаимосвязи нарушений функционального статуса симпатической или парасимпатической нервной системы при воспалении — важнейшие направления научного поиска, позволяющие перспективно оценивать возможности современной интенсивной терапии.
Вклад авторов: Ю.Ю. Кирячков — концепция статьи и систематизация современных знаний по теме обзора; М.В. Петрова — дизайн статьи; С.А. Босенко — перевод используемых научных статьей; Б.Г. Муслимов — интерпретация данных, относящихся непосредственно к клиническим исследованиям.
Источник финансирования. Работа выполнена за счет средств бюджета Федерального научно-клинического центра реаниматологии и реабилитологии.
Конфликт интересов. Авторы заявляют об отсутствии конфликта интересов.
Литература
- Williams D.P., Koenig J., Carnevali L., Sgoifo A., Jarczok M.N., Sternberg E.M., Thayer J.F. Heart rate variability and inflammation: a meta-analysis of human studies. Brain Behav Immun 2019; 80: 219–226, https://doi.org/10.1016/j.bbi.2019.03.009.
- Werdan K., Schmidt H., Ebelt H., Zorn-Pauly K., Koidl B., Hoke R.S., Heinroth K., Müller-Werdan U. Impaired regulation of cardiac function in sepsis, SIRS, and MODS. Can J Physiol Pharmacol 2009; 87(4): 266–274, https://doi.org/10.1139/Y09-012.
- Raithel D.S., Ohler K.H., Porto I., Bicknese A.R., Kraus D.M. Morphine: an effective abortive therapy for pediatric paroxysmal sympathetic hyperactivity after hypoxic brain injury. J Pediatr Pharmacol Ther 2015; 20(4): 335–340.
- Baguley I.J., Perkes I.E., Fernandez-Ortega J.F., Rabinstein A.A., Dolce G., Hendricks H.T.; Consensus Working Group. Paroxysmal sympathetic hyperactivity after acquired brain injury: consensus on conceptual definition, nomenclature, and diagnostic criteria. J Neurotrauma 2014; 31(17): 1515–1520, https://doi.org/10.1089/neu.2013.3301.
- Esterov D., Greenwald B.D. Autonomic dysfunction after mild traumatic brain injury. Brain Sci 2017; 7(8): E100, https://doi.org/10.3390/brainsci7080100.
- Wang D.W., Yin Y.M., Yao Y.M. Vagal modulation of the inflammatory response in sepsis. Int Rev Immunol 2016; 35(5): 415–433, https://doi.org/10.3109/08830185.2015.1127369.
- Zila I., Mokra D., Kopincova J., Kolomaznik M., Javorka M., Calkovska A. Vagal-immune interactions involved in cholinergic anti-inflammatory pathway. Physiol Res 2017; 66(Suppl 2): S139–S145, https://doi.org/10.33549/physiolres.933671.
- Huang Y., Zhao C., Su X. Neuroimmune regulation of lung infection and inflammation. QJM 2019; 112(7): 483–487, https://doi.org/10.1093/qjmed/hcy154.
- Ren C., Li X.H., Wang S.B., Wang L.X., Dong N., Wu Y., Yao Y.M. Activation of central alpha 7 nicotinic acetylcholine receptor reverses suppressed immune function of T lymphocytes and protects against sepsis lethality. Int J Biol Sci 2018; 14(7): 748–759, https://doi.org/10.7150/ijbs.24576.
- Reyes-Lagos J.J., Ledesma-Ramírez C.I., Pliego-Carrillo A.C., Peña-Castillo M.Á., Echeverría J.C., Becerril-Villanueva E., Pavón L., Pacheco-López G. Neuroautonomic activity evidences parturition as a complex and integrated neuro-immune-endocrine process. Ann N Y Acad Sci 2018; 1437(1): 22–30, https://doi.org/10.1111/nyas.13860.
- Qian Y.S., Zhao Q.Y., Zhang S.J., Zhang Y.J., Wang Y.C., Zhao H.Y., Dai Z.X., Tang Y.H., Wang X., Wang T., Huang C.X. Effect of α7nAChR mediated cholinergic anti-inflammatory pathway on inhibition of atrial fibrillation by low-level vagus nerve stimulation. Zhonghua Yi Xue Za Zhi 2018; 98(11): 855–859, https://doi.org/10.3760/cma.j.issn.0376-2491.2018.11.013.
- Murray K., Reardon C. The cholinergic anti-inflammatory pathway revisited. Neurogastroenterol Motil 2018; 30(3), https://doi.org/10.1111/nmo.13288.
- Yamada M., Ichinose M. The cholinergic anti-inflammatory pathway: an innovative treatment strategy for respiratory diseases and their comorbidities. Curr Opin Pharmacol 2018; 40: 18–25, https://doi.org/10.1016/j.coph.2017.12.003.
- Pavlov V.A., Ochani M., Yang L.H., Gallowitsch-Puerta M., Ochani K., Lin X., Levi J., Parrish W.R., Rosas-Ballina M., Czura C.J., Larosa G.J., Miller E.J., Tracey K.J., Al-Abed Y. Selective α7-nicotinic acetylcholine receptor agonist GTS-21 improves survival in murine endotoxemia and severe sepsis. Crit Care Med 2007; 35(4): 1139–1144, https://doi.org/10.1097/01.CCM.0000259381.56526.96.
- Lu J., Goh S.J., Tng P.Y., Deng Y.Y., Ling E.A., Moochhala S. Systemic inflammatory response following acute traumatic brain injury. Front Biosci (Landmark Ed) 2009; 14: 3795–3813, https://doi.org/10.2741/3489.
- Shin S.S., Dixon C.E. Alterations in cholinergic pathways and therapeutic strategies targeting cholinergic system after traumatic brain injury. J Neurotrauma 2015; 32(19): 1429–1440, https://doi.org/10.1089/neu.2014.3445.
- Liu Q., Xie J., Yang Y. Advances in the regulation mechanism of cholinergic anti-inflammatory pathway on sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2019; 31(6): 781–784, https://doi.org/10.3760/cma.j.issn.2095-4352.2019.06.024.
- Guyot M., Simon T., Panzolini C., Ceppo F., Daoudlarian D., Murris E., Macia E., Abélanet S., Sridhar A., Vervoordeldonk M.J., Glaichenhaus N., Blancou P. Apical splenic nerve electrical stimulation discloses an anti-inflammatory pathway relying on adrenergic and nicotinic receptors in myeloid cells. Brain Behav Immun 2019; 80: 238–246, https://doi.org/10.1016/j.bbi.2019.03.015.
- Noh H., Yu M.R., Kim H.J., Lee J.H., Park B.W., Wu I., Matsumoto M., King G.L. Beta 2 adrenergic receptor agonists are novel regulators of macrophage activation in diabetic renal and cardiovascular complications. Kidney Int 2017; 92(1): 101–113, https://doi.org/10.1016/j.kint.2017.02.013.
- Lechtenberg K.J., Meyer S.T., Doyle J.B., Peterson T.C., Buckwalter M.S. Augmented β2-adrenergic signaling dampens the neuroinflammatory response following ischemic stroke and increases stroke size. J Neuroinflammation 2019; 16(1): 112, https://doi.org/10.1186/s12974-019-1506-4.
- Zabrodskii P.F., Gromov M.S., Maslyakov V.V. Combined effect of NF-κB inhibitor and β2-adrenoreceptor agonist on mouse mortality and blood concentration of proinflammatory cytokines in sepsis. Bull Exp Biol Med 2018; 165(4): 445–448, https://doi.org/10.1007/s10517-018-4190-y.
- Sun J.J., Lan J.F., Zhao X.F., Vasta G.R., Wang J.X. Binding of a C-type lectin’s coiled-coil domain to the Domeless receptor directly activates the JAK/STAT pathway in the shrimp immune response to bacterial infection. PLoS Pathog 2017; 13(9): e1006626, https://doi.org/10.1371/journal.ppat.1006626.
- Ruiz-Medina B.E., Cadena-Medina D.A., Esparza E., Arrieta A.J., Kirken R.A. Isoproterenol-induced beta-2 adrenergic receptor activation negatively regulates interleukin-2 signaling. Biochem J 2018; 475(18): 2907–2923, https://doi.org/10.1042/BCJ20180503.
- Wu H., Li L., Su X. Vagus nerve through α7 nAChR modulates lung infection and inflammation: models, cells, and signals. Biomed Res Int 2014; 2014: 283525, https://doi.org/10.1155/2014/283525.
- Vaughn A.C., Cooper E.M., DiLorenzo P.M., O’Loughlin L.J., Konkel M.E., Peters J.H., Hajnal A., Sen T., Lee S.H., de La Serre C.B., Czaja K. Energy-dense diet triggers changes in gut microbiota, reorganization of gut brain vagal communication and increases body fat accumulation. Acta Neurobiol Exp (Wars) 2017; 77(1): 18–30, https://doi.org/10.21307/ane-2017-033.
- Cawthon C.R., de La Serre C.B. Gut bacteria interaction with vagal afferents. Brain Res 2018; 1693 (Pt B): 134–139, https://doi.org/10.1016/j.brainres.2018.01.012.
- Bonaz B., Sinniger V., Pellissier S. Vagus nerve stimulation at the interface of brain-gut interactions. Cold Spring Harb Perspect Med 2019; 9(8): a034199, https://doi.org/10.1101/cshperspect.a034199.
- Houlden A., Goldrick M., Brough D., Vizi E.S., Lenart N., Martinecz B., Roberts I.S., Denes A. Brain injury induces specific changes in the caecal microbiota of mice via altered autonomic activity and mucoprotein production. Brain Behav Immun 2016; 57: 10–20, https://doi.org/10.1016/j.bbi.2016.04.003.
- Kigerl K.A., Mostacada K., Popovich P.G. Gut microbiota are disease-modifying factors after traumatic spinal cord injury. Neurotherapeutics 2018; 15(1): 60–67, https://doi.org/10.1007/s13311-017-0583-2.
- de Jonge W.J. The gut’s little brain in control of intestinal immunity. ISRN Gastroenterol 2013; 2013: 630159, https://doi.org/10.1155/2013/630159.
- Radmark L., Sidorchuk A., Osika W., Niemi M. A systematic review and meta-analysis of the impact of mindfulness based interventions on heart rate variability and inflammatory markers. J Clin Med 2019; 8(10): E1638, https://doi.org/10.3390/jcm8101638.
- Sharma V.K., Savitha S., Vinod K.V., Rajappa M., Subramanian S.K., Rajendran R. Assessment of autonomic functions and its association with telomerase level, oxidative stress and inflammation in complete glycemic spectrum — an exploratory study. Diabetes Metab Syndr 2019; 13(2): 1193–1199, https://doi.org/10.1016/j.dsx.2019.01.011.
- Pong J.Z., Fook-Chong S., Koh Z.X., Samsudin M.I., Tagami T., Chiew C.J., Wong T.H., Ho A.F.W., Ong M.E.H., Liu N. Combining heart rate variability with disease severity score variables for mortality risk stratification in septic patients presenting at the emergency department. Int J Environ Res Public Health 2019; 16(10): E1725, https://doi.org/10.3390/ijerph16101725.
- Rupprecht S., Finn S., Hoyer D., Guenther A., Witte O.W., Schultze T., Schwab M. Association between systemic inflammation, carotid arteriosclerosis, and autonomic dysfunction. Transl Stroke Res 2020; 11(1): 50–59, https://doi.org/10.1007/s12975-019-00706-x.
- Magrone T., Jirillo E. Sepsis: from historical aspects to novel vistas. pathogenic and therapeutic considerations. Endocr Metab Immune Disord Drug Targets 2019; 19(4): 490–502, https://doi.org/10.2174/1871530319666181129112708.
- Kanashiro A., Sônego F., Ferreira R.G., Castanheira F.V., Leite C.A., Borges V.F., Nascimento D.C., Cólon D.F., Alves-Filho J.C., Ulloa L., Cunha F.Q. Therapeutic potential and limitations of cholinergic anti-inflammatory pathway in sepsis. Pharmacol Res 2017; 117: 1–8, https://doi.org/10.1016/j.phrs.2016.12.014.
- Assinger A., Schrottmaier W.C., Salzmann M., Rayes J. Platelets in sepsis: an update on experimental models and clinical data. Front Immunol 2019; 10: 1687, https://doi.org/10.3389/fimmu.2019.01687.
- Vergadi E., Vaporidi K., Tsatsanis C. Regulation of endotoxin tolerance and compensatory anti-inflammatory response syndrome by non-coding RNAs. Front Immunol 2018; 9: 2705, https://doi.org/10.3389/fimmu.2018.02705.
- Quek A.M.L., Britton J.W., McKeon A., So E., Lennon V.A., Shin C., Klein C., Watson R.E., Kotsenas A.L., Lagerlund T.D., Cascino G.D., Worrell G.A., Wirrell E.C., Nickels K.C., Aksamit A.J., Noe K.H., Pittock S.J. Autoimmunne epilepsy: clinical characteristics and response to immunotherapy. Arch Neurol 2012; 69(5): 582–593, https://doi.org/10.1001/archneurol.2011.2985.
- Bauer J., Becker A.J., Elyaman W., Peltola J., Rüegg S., Titulaer M.J., Varley J.A., Beghi E. Innate and adaptive immunity in human epilepsies. Epilepsia 2017; 58(Suppl 3): 57–68, https://doi.org/10.1111/epi.13784.
- Golden E.P., Vernino S. Autoimmune autonomic neuropathies and ganglionopathies: epidemiology, pathophysiology, and therapeutic advances. Clin Auton Res 2019; 29(3): 277–288, https://doi.org/10.1007/s10286-019-00611-1.
- Gaddam S.S., Buell T., Robertson C.S. Systemic manifestations of traumatic brain injury. Handb Clin Neurol 2015; 127: 205–218, https://doi.org/10.1016/B978-0-444-52892-6.00014-3.
- Frasch M.G., Szynkaruk M., Prout A.P., Nygard K., Cao M., Veldhuizen R., Hammond R., Richardson B.S. Decreased neuroinflammation correlates to higher vagus nerve activity fluctuations in near-term ovine fetuses: a case for the afferent cholinergic anti-inflammatory pathway? J Neuroinflammation 2016; 13(1): 103, https://doi.org/10.1186/s12974-016-0567-x.
- Nicholls A.J., Wen S.W., Hall P., Hickey M.J., Wong C.H.Y. Activation of the sympathetic nervous system modulates neutrophil function. J Leukoc Biol 2018; 103(2): 295–309, https://doi.org/10.1002/JLB.3MA0517-194RR.
- Oikawa S., Kai Y., Mano A., Sugama S., Mizoguchi N., Tsuda M., Muramoto K., Kakinuma Y. Potentiating a non-neuronal cardiac cholinergic system reinforces the functional integrity of the blood brain barrier associated with systemic anti-inflammatory responses. Brain Behav Immun 2019; 81: 122–137, https://doi.org/10.1016/j.bbi.2019.06.005.
- Boeckxstaens G. The clinical importance of the anti-inflammatory vagovagal reflex. Handb Clin Neurol 2013; 117: 119–134, https://doi.org/10.1016/b978-0-444-53491-0.00011-0.
- Nunes N.S., Chandran P., Sundby M., Visioli F., da Costa Gonçalves F., Burks S.R., Paz A.H., Frank J.A. Therapeutic ultrasound attenuates DSS-induced colitis through the cholinergic anti-inflammatory pathway. EBioMedicine 2019; 45: 495–510, https://doi.org/10.1016/j.ebiom.2019.06.033.
- Inoue T., Abe C., Kohro T., Tanaka S., Huang L., Yao J., Zheng S., Ye H., Inagi R., Stornetta R.L., Rosin D.L., Nangaku M., Wada Y., Okusa M.D. Non-canonical cholinergic anti-inflammatory pathway-mediated activation of peritoneal macrophages induces Hes1 and blocks ischemia/reperfusion injury in the kidney. Kidney Int 2019; 95(3): 563–576, https://doi.org/10.1016/j.kint.2018.09.020.
- Li Z., Hao H., Gao Y., Wang Z., Lu W., Liu J. Expression and localization analyses of the cholinergic anti-inflammatory pathway and α7nAchR in different tissues of rats with rheumatoid arthritis. Acta Histochem 2019; 121(6): 742–749, https://doi.org/10.1016/j.acthis.2019.07.001.
- Yamada M., Ichinose M. The cholinergic pathways in inflammation: a potential pharmacotherapeutic target for COPD. Front Pharmacol 2018; 9: 1426, https://doi.org/10.3389/fphar.2018.01426.
- Tian Y., Miao B., Charles E.J., Wu D., Kron I.L., French B.A., Yang Z. Stimulation of the beta2 adrenergic receptor at reperfusion limits myocardial reperfusion injury via an interleukin-10-dependent anti-inflammatory pathway in the spleen. Circ J 2018; 82(11): 2829–2836, https://doi.org/10.1253/circj.CJ-18-0061.
- Antunes G.L., Silveira J.S., Kaiber D.B., Luft C., da Costa M.S., Marques E.P., Ferreira F.S., Breda R.V., Wyse A.T.S., Stein R.T., Pitrez P.M., da Cunha A.A. Cholinergic anti-inflammatory pathway confers airway protection against oxidative damage and attenuates inflammation in an allergic asthma model. J Cell Physiol 2020; 235(2): 1838–1849, https://doi.org/10.1002/jcp.29101.
- Jarczyk J., Yard B.A., Hoeger S. The cholinergic anti-inflammatory pathway as a conceptual framework to treat inflammation-mediated renal injury. Kidney Blood Press Res 2019; 44(4): 435–448, https://doi.org/10.1159/000500920.
- Hajiasgharzadeh K., Baradaran B. Cholinergic anti-inflammatory pathway and the liver. Adv Pharm Bull 2017; 7(4): 507–513, https://doi.org/10.15171/apb.2017.063.
- Xu H., Shi Q., Mo Y., Wu L., Gu J., Xu Y. Downregulation of α7 nicotinic acetylcholine receptors in peripheral blood monocytes is associated with enhanced inflammation in preeclampsia. BMC Pregnancy Childbirth 2019; 19(1): 188, https://doi.org/10.1186/s12884-019-2340-5.
- Fernández-Cabezudo M.J., George J.A., Bashir G., Mohamed Y.A., Al-Mansori A., Qureshi M.M., Lorke D.E., Petroianu G., Al-Ramadi B.K. Involvement of acetylcholine receptors in cholinergic pathway-mediated protection against autoimmune diabetes. Front Immunol 2019; 10: 1038, https://doi.org/10.3389/fimmu.2019.01038.
- Gálvez I., Martín-Cordero L., Hinchado M.D., Álvarez-Barrientos A., Ortega E. Anti-inflammatory effect of β2 adrenergic stimulation on circulating monocytes with a pro-inflammatory state in high-fat diet-induced obesity. Brain Behav Immun 2019; 80: 564–572, https://doi.org/10.1016/j.bbi.2019.04.042.
- Ortega E., Gálvez I., Martín-Cordero L. Adrenergic regulation of macrophage-mediated innate/inflammatory responses in obesity and exercise in this condition: role of β2 adrenergic receptors. Endocr Metab Immune Disord Drug Targets 2019; 19(8): 1089–1099, https://doi.org/10.2174/1871530319666190206124520.
- Hatakeyama N., Matsuda N. Alert cell strategy: mechanisms of inflammatory response and organ protection. Curr Pharm Des 2014; 20(36): 5766–5778, https://doi.org/10.2174/138161282036140912122809.
- Clar D.T., Sharma S. Autonomic pharmacology. In: StatPearls. Treasure Island (FL): StatPearls Publishing; 2019. URL: https://www.ncbi.nlm.nih.gov/books/NBK538180.
- Samuel S., Allison T.A., Lee K., Choi H.A. Pharmacologic management of paroxysmal sympathetic hyperactivity after brain injury. J Neurosci Nurs 2016; 48(2): 82–89, https://doi.org/10.1097/JNN.0000000000000207.
- Lehner K.R., Silverman H.A., Addorisio M.E., Roy A., Al-Onaizi M.A., Levine Y., Olofsson P.S., Chavan S.S., Gros R., Nathanson N.M., Al-Abed Y., Metz C.N., Prado V.F., Prado M.A.M., Tracey K.J., Pavlov V.A. Forebrain cholinergic signaling regulates innate immune responses and inflammation. Front Immunol 2019; 10: 585, https://doi.org/10.3389/fimmu.2019.00585.
- Chang E.H., Chavan S.S., Pavlov V.A. Cholinergic control of inflammation, metabolic dysfunction, and cognitive impairment in obesity-associated disorders: mechanisms and novel therapeutic opportunities. Front Neurosci 2019; 13: 263, https://doi.org/10.3389/fnins.2019.00263.
- Njoku I., Radabaugh H.L., Nicholas M.A., Kutash L.A., O’Neil D.A., Marshall I.P., Cheng J.P., Kline A.E., Bondi C.O. Chronic treatment with galantamine rescues reversal learning in an attentional set-shifting test after experimental brain trauma. Exp Neurol 2019; 315: 32–41, https://doi.org/10.1016/j.expneurol.2019.01.019.
- Pinder N., Bruckner T., Lehmann M., Motsch J., Brenner T., Larmann J., Knebel P., Hoppe-Tichy T., Swoboda S., Weigand M.A., Hofer S., Zimmermann J.B. Effect of physostigmine on recovery from septic shock following intra-abdominal infection — results from a randomized, double-blind, placebo-controlled, monocentric pilot trial (Anticholium® per Se). J Crit Care 2019; 52: 126–135, https://doi.org/10.1016/j.jcrc.2019.04.012.
- Chen Y., Zhang X., Zhang B., He G., Zhou L., Xie Y. Dexmedetomidine reduces the neuronal apoptosis related to cardiopulmonary bypass by inhibiting activation of the JAK2-STAT3 pathway. Drug Des Devel Ther 2017; 11: 2787–2799, https://doi.org/10.2147/DDDT.S140644.
- Xu K.L., Liu X.Q., Yao Y.L., Ye M.R., Han Y.G., Zhang T., Chen G., Lei M. Effect of dexmedetomidine on rats with convulsive status epilepticus and association with activation of cholinergic anti-inflammatory pathway. Biochem Biophys Res Commun 2018; 495(1): 421–426, https://doi.org/10.1016/j.bbrc.2017.10.124.
- Yamanaka D., Kawano T., Nishigaki A., Aoyama B., Tateiwa H., Shigematsu-Locatelli M., Locatelli F.M., Yokoyama M. Preventive effects of dexmedetomidine on the development of cognitive dysfunction following systemic inflammation in aged rats. J Anesth 2017; 31(1): 25–35, https://doi.org/10.1007/s00540-016-2264-4.
- Cai Y., Xu H., Yan J., Zhang L., Lu Y. Molecular targets and mechanism of action of dexmedetomidine in treatment of ischemia/reperfusion injury. Mol Med Rep 2014; 9(5): 1542–1550, https://doi.org/10.3892/mmr.2014.2034.
- Zhang J., Xia F., Zhao H., Peng K., Liu H., Meng X., Chen C., Ji F. Dexmedetomidine-induced cardioprotection is mediated by inhibition of high mobility group box-1 and the cholinergic anti-inflammatory pathway in myocardial ischemia-reperfusion injury. PLoS One 2019; 14(7): e0218726, https://doi.org/10.1371/journal.pone.0218726.
- Jiang L., Hu M., Lu Y., Cao Y., Chang Y., Dai Z. The protective effects of dexmedetomidine on ischemic brain injury: a meta-analysis. J Clin Anesth 2017; 40: 25–32, https://doi.org/10.1016/j.jclinane.2017.04.003.
- Hu J., Vacas S., Feng X., Lutrin D., Uchida Y., Lai I.K., Maze M. Dexmedetomidine prevents cognitive decline by enhancing resolution of high mobility group box 1 protein-induced inflammation through a vagomimetic action in mice. Anesthesiology 2018; 128(5): 921–931, https://doi.org/10.1097/ALN.0000000000002038.
- Janssen T.L., Alberts A.R., Hooft L., Mattace-Raso F., Mosk C.A., van der Laan L. Prevention of postoperative delirium in elderly patients planned for elective surgery: systematic review and meta-analysis. Clin Interv Aging 2019; 14: 1095–1117, https://doi.org/10.2147/CIA.S201323.
- Lankadeva Y.R., Ma S., Iguchi N., Evans R.G., Hood S.G., Farmer D.G.S., Bailey S.R., Bellomo R., May C.N. Dexmedetomidine reduces norepinephrine requirements and preserves renal oxygenation and function in ovine septic acute kidney injury. Kidney Int 2019; 96(5): 1150–1161, https://doi.org/10.1016/j.kint.2019.06.013.
- Zi S.F., Li J.H., Liu L., Deng C., Ao X., Chen D.D., Wu S.Z. Dexmedetomidine-mediated protection against septic liver injury depends on TLR4/MyD88/NF-κB signaling downregulation partly via cholinergic anti-inflammatory mechanisms. Int Immunopharmacol 2019; 76: 105898, https://doi.org/10.1016/j.intimp.2019.105898.
- Suzuki T., Suzuki Y., Okuda J., Kurazumi T., Suhara T., Ueda T., Nagata H., Morisaki H. Sepsis-induced cardiac dysfunction and β-adrenergic blockade therapy for sepsis. J Intensive Care 2017; 5: 22, https://doi.org/10.1186/s40560-017-0215-2.
- Brown S.M., Beesley S.J., Lanspa M.J., Grissom C.K., Wilson E.L., Parikh S.M., Sarge T., Talmor D., Banner-Goodspeed V., Novack V., Thompson B.T., Shahul S.; Esmolol to Control Adrenergic Storm in Septic Shock-ROLL-IN (ECASSS-R) study. Esmolol infusion in patients with septic shock and tachycardia: a prospective, single-arm, feasibility study. Pilot Feasibility Study 2018; 4: 132, https://doi.org/10.1186/s40814-018-0321-5.
- Breit S., Kupferberg A., Rogler G., Hasler G. Vagus nerve as modulator of the brain-gut axis in psychiatric and inflammatory disorders. Front Psychiatry 2018; 9: 44, https://doi.org/10.3389/fpsyt.2018.00044.
- Carod-Artal F.J. Infectious diseases causing autonomic dysfunction. Clin Auton Res 2017; 28(1): 67–81, https://doi.org/10.1007/s10286-017-0452-4.
- Han C., Rice M.W., Cai D. Neuroinflammatory and autonomic mechanisms in diabetes and hypertension. Am J Physiol Endocrinol Metab 2016; 311(1): E32–E41, https://doi.org/10.1152/ajpendo.00012.2016.
- Gao H., Molinas A.J.R., Miyata K., Qiao X., Zsombok A. Overactivity of liver-related neurons in the paraventricular nucleus of the hypothalamus: electrophysiological findings in db/db mice. J Neurosci 2017; 37(46): 11140–11150, https://doi.org/10.1523/JNEUROSCI.1706-17.2017.
- Hong G.S., Zillekens A., Schneiker B., Pantelis D., de Jonge W.J., Schaefer N., Kalff J.C., Wehner S. Non-invasive transcutaneous auricular vagus nerve stimulation prevents postoperative ileus and endotoxemia in mice. Neurogastroenterol Motil 2019; 31(3): e13501, https://doi.org/10.1111/nmo.13501.
- Bosmans G., Appeltans I., Stakenborg N., Gomez-Pinilla P.J., Florens M.V., Aguilera-Lizarraga J., Matteoli G., Boeckxstaens G.E. Vagus nerve stimulation dampens intestinal inflammation in a murine model of experimental food allergy. Allergy 2019; 74(9): 1748–1759, https://doi.org/10.1111/all.13790.
- Papaioannou V., Pnevmatikos I. Heart rate variability: a potential tool for monitoring immunomodulatory effects of parenteral fish oil feeding in patients with sepsis. Nutr Metab Insights 2019; 12: 1178638819847486, https://doi.org/10.1177/1178638819847486.
- Hoover В.B. Cholinergic modulation of the immune system presents new approaches for treating inflammation. Pharmacol Ther 2017; 179: 1–16, https://doi.org/10.1016/j.pharmthera.2017.05.002.
- Tao G., Min-Hua C., Feng-Chan X., Yan C., Ting S., Wei-Qin L., Wen-Kui Y. Changes of plasma acetylcholine and inflammatory markers in critically ill patients during early enteral nutrition: a prospective observational study. J Crit Care 2019; 52: 219–226, https://doi.org/10.1016/j.jcrc.2019.05.008.
- Roewe J., Higer M., Riehl D.R., Gericke A., Radsak M.P., Bosmann M. Neuroendocrine modulation of IL-27 in macrophages. J Immunol 2017; 199(7): 2503–2514, https://doi.org/10.4049/jimmunol.1700687.
- Ağaç D., Estrada L.D., Maples R., Hooper L.V., Farrar J.D. The β2-adrenergic receptor controls inflammation by driving rapid IL-10 secretion. Brain Behav Immun 2018; 74: 176–185, https://doi.org/10.1016/j.bbi.2018.09.004.
- Li H.M., Li K.Y., Xing Y., Tang X.X., Yang D.M., Dai X.M., Lu D.X., Wang H.D. Phenylephrine attenuated sepsis-induced cardiac inflammation and mitochondrial injury through an effect on the PI3K/Akt signaling pathway. J Cardiovasc Pharmacol 2019; 73(3): 186–194, https://doi.org/10.1097/FJC.0000000000000651.
- Mogilevski T., Burgell R., Aziz Q., Gibson P.R. Review article: the role of the autonomic nervous system in the pathogenesis and therapy of IBD. Aliment Pharmacol Ther 2019; 50(7): 720–737, https://doi.org/10.1111/apt.15433.
- Sonnenberg G.F., Hepworth M.R. Functional interactions between innate lymphoid cells and adaptive immunity. Nat Rev Immunol 2019; 19(10): 599–613, https://doi.org/10.1038/s41577-019-0194-8.
- Serhan C.N., de la Rosa X., Jouvene C. Novel mediators and mechanisms in the resolution of infectious inflammation: evidence for vagus regulation. J Intern Med 2019; 286(3): 240–258, https://doi.org/10.1111/joim.12871.
- Carnagarin R., Matthews V., Zaldivia M.T.K., Schlaich M.P. The bidirectional interaction between the sympathetic nervous system and immune mechanisms in the pathogenesis of hypertension. Br J Pharmacol 2019; 176(12): 1839–1852, https://doi.org/10.1111/bph.14481.
- Reardon C. Neuro-immune interactions in the cholinergic anti-inflammatory reflex. Immunol Lett 2016; 178: 92–96, https://doi.org/10.1016/j.imlet.2016.08.006.
- Eduardo C.-R.C., Alejandra T.-I.G., Guadalupe D.-R.K.J., Herminia V.-R.G., Lenin P., Enrique B.-V., Evandro B.M., Oscar B., Iván G.-P.M. Modulation of the extraneuronal cholinergic system on main innate response leukocytes. J Neuroimmunol 2019; 327: 22–35, https://doi.org/10.1016/j.jneuroim.2019.01.008.
- Pereira M.R., Leite P.E. The involvement of parasympathetic and sympathetic nerve in the inflammatory reflex. J Cell Physiol 2016; 231(9): 1862–1869, https://doi.org/10.1002/jcp.25307.
- Kim H.G., Cheon E.J., Bai D.S., Lee Y.H., Koo B.H. Stress and heart rate variability: a meta-analysis and review of the literature. Psychiatry Investig 2018; 15(3): 235–245, https://doi.org/10.30773/pi.2017.08.17.
- Liu L., Zhao M., Yu X., Zang W. Pharmacological modulation of vagal nerve activity in cardiovascular diseases. Neurosci Bull 2019; 35(1): 156–166, https://doi.org/10.1007/s12264-018-0286-7.
- Crippa I.A., Subirà C., Vincent J.L., Fernandez R.F., Hernandez S.C., Cavicchi F.Z., Creteur J., Taccone F.S. Impaired cerebral autoregulation is associated with brain dysfunction in patients with sepsis. Crit Care 2018; 22(1): 327, https://doi.org/10.1186/s13054-018-2258-8.
- Sanz D., D’Arco F., Robles C.A., Brierley J. Incidence and pattern of brain lesions in paediatric septic shock patients. Br J Radiol 2018; 91(1084): 20170861, https://doi.org/10.1259/bjr.20170861.
- Esen F., Orhun G., Özcan P.E., Brenes Bastos A.R., Tüzün E. Diagnosing acute brain dysfunction due to sepsis. Neurol Sci 2020; 41(1): 25–33, https://doi.org/10.1007/s10072-019-04069-x.
- Günther A., Schubert J., Witte O.W., Brämer D. Intensive care aspects of autoimmune encephalitis. Med Klin Intensivmed Notfmed 2019; 14(7): 620–627, https://doi.org/10.1007/s00063-019-0604-5.
- Bonjorno Junior J.C., Caruso F.R., Mendes R.G., da Silva T.R., Biazon T.M.P.C., Rangel F., Phillips S.A., Arena R., Borghi-Silva A. Noninvasive measurements of hemodynamic, autonomic and endothelial function as predictors of mortality in sepsis: a prospective cohort study. PLoS One 2019; 14(3): e0213239, https://doi.org/10.1371/journal.pone.0213239.
- Biteker F.S., Özlek B., Çelik O., Özlek E., Çil C., Doğan V., Biteker M. Autonomic imbalance in sepsis. Am J Emerg Med 2018; 36(2): 322, https://doi.org/10.1016/j.ajem.2017.11.054.
- Tracey K.J. The inflammatory reflex. Nature 2002; 420(6917): 853–859, https://doi.org/10.1038/nature01321.








